
eBook - ePub
Local Anaesthesia in Dentistry
J. A. Baart, H. S. Brand
This is a test
Share book
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Local Anaesthesia in Dentistry
J. A. Baart, H. S. Brand
Book details
Book preview
Table of contents
Citations
About This Book
Local Anaesthesia in Dentistry is a practical guide for both students and general practitioners to this essential area of clinical practice.
Highly illustrated in full colour throughout, the book provides clear and practical guidance to the administration of local anaesthesia. The book introduces the reader to the concept of nerve conduction and pain as well as providing an explanation of the anatomy of the trigeminal nerve. Further chapters cover such key areas of practice as regional anaesthesia, local anaesthesia in children, pharmacology, local and systemic complications, general practical aspects, the prevention of side effects and legal aspects.
Frequently asked questions
How do I cancel my subscription?
Can/how do I download books?
At the moment all of our mobile-responsive ePub books are available to download via the app. Most of our PDFs are also available to download and we're working on making the final remaining ones downloadable now. Learn more here.
What is the difference between the pricing plans?
Both plans give you full access to the library and all of Perlego’s features. The only differences are the price and subscription period: With the annual plan you’ll save around 30% compared to 12 months on the monthly plan.
What is Perlego?
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 1000+ topics, we’ve got you covered! Learn more here.
Do you support text-to-speech?
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more here.
Is Local Anaesthesia in Dentistry an online PDF/ePUB?
Yes, you can access Local Anaesthesia in Dentistry by J. A. Baart, H. S. Brand in PDF and/or ePUB format, as well as other popular books in Medicine & Dentistry. We have over one million books available in our catalogue for you to explore.
Information
1
Pain and impulse conduction
According to the World Health Organisation pain is defined as an ‘unpleasant sensation that occurs from imminent tissue damage’. From a physiological perspective, pain is a warning system. During dental treatment, patients will experience pain as something unpleasant.
1.1 Pain receptors
Pain stimuli are primarily generated by the relatively amorph sensory nerve endings of the Aδ and C fibres. These free nerve endings (nociceptors; see Figure 1.1) are sensitive to a variety of mechanical, thermal and chemical stimuli and are therefore called polymodal. Nociceptors do not display adaptation: nociceptive responses will occur as long as the stimulus is present. Nociceptors have a high threshold for activation. The detection of the stimulus is performed by the receptors, present on the sensory nerve endings, that convert the stimulus into an electric signal. This process is called transduction.
During tissue damage, several substances are released that are able to stimulate the nociceptors, such as histamine, serotonin, bradykinin, prostaglandin E2 and interleukins. These substances activate the nociceptors and reduce their threshold (sensitisation).
Nociceptors are also present in the teeth and the oral cavity and are usually sensitive to a specific neurotransmitter. Most nerve fibres, however, contain various nociceptors.
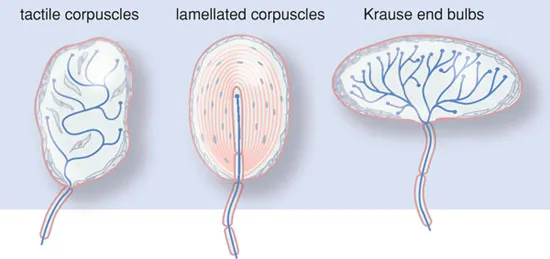
The sensory nervous system also contains ‘physiological’ sensors. These are small end organs of the sensory nerves, such as the Krause, Meisner and Pacini bodies (see Figure 1.2). These ‘physiological’ sensors usually only respond to one specific stimulus (warmth, touch, smell, etc.) and are, as such, unimodal. Besides this, they exhibit the phenomenon of adaptation; the response to stimulus disappears during prolonged or persistent stimulation. In the case of excessive stimulation, these ‘physiological’ sensors may also initiate pain sensation.
Figure 1.1 Nociceptors.
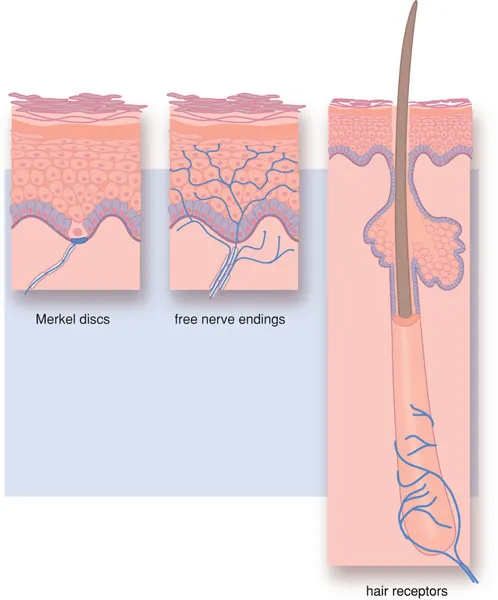
Figure 1.2 Physiological sensors.
1.2 Nerve impulse transmission
The stimuli, received by the nociceptors and converted into nerve impulses, eventually must be interpreted in the brain. The nerve impulse is transported within the sensory nervous system, wherein three nerve fibres are successively linked. The first nerve fibres form the peripheral nerve. The second and third are present in the central nervous system and form nerve bundles (pathways or tracts). The cell nuclei of the individual neurons are grouped together in ganglia and nuclei.
1.2.1 The structure of the peripheral nerve
Nociceptive stimuli are transported along sensory thinly myelinated Aδ and unmyelinated C fibres. Other types of nerve fibres are involved in the transport of other sensory stimuli (see Box 1.1).
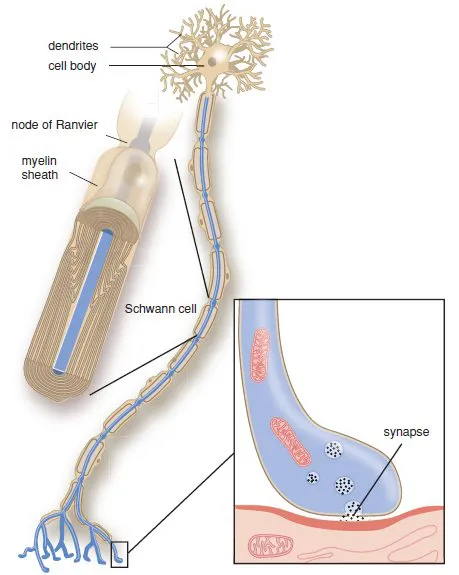
A peripheral nerve is composed of nerve fibres from a group of neurons, enwrapped in a connective tissue network. The individual fibres may, or may not be, surrounded by an isolating myelin layer, Schwann’s sheath. The cell body is the metabolic centre of the neuron (Figure 1.3) where most cell organelles are produced. Dendrites transport impulses towards the cell body and axons transmit signals away from the cell body. Some axons are surrounded by a myelin sheath, others are not. The axons and dendrites are elongated and form the nerve fibres. At the end of the dendrites, receptors are present that can receive signals. At the end of the axons are synapses, where the impulse is transmitted to another nerve cell or to a cell of the end organ.
Box 1.1 Nociceptive pathways
In the body, nociceptive stimuli are received by nociceptors and then propagated via an Aδ or C fibre. The first are thinly myelinated with a fast transmission of stimuli, whereas the second are unmyelinated with a slow transmission.
The C fibres conduct impulses generated by temperature, pain and itching. The Aα fibres conduct motor impulses for the body’s posture and movement; the Aβ fibres transport impulses generated by touch and signals from the skin mechano-receptors; and the Aδ fibres conduct pain impulses, temperature signals and signals to maintain the muscular tone.
The cell bodies of these primary neurons are located in the dorsal root ganglion. The axons run through Lissauer’s tract to the dorsal horn of the spinal cord, where they connect to the secondary sensory neuron in Rexed’s laminae. This secondary sensory neuron crosses the midline and ascends as the spinothalamic tract. The spinothalamic tract forms synapses with nuclei of the thalamus, where it projects onto the somatosensory cortex. Descending pathways from the somatosensory cortex modulate the nociceptive system. From these fibres, the neurotransmitters serotonin and noradrenalin are released.
Figure B1.1 Primary afferent axons.
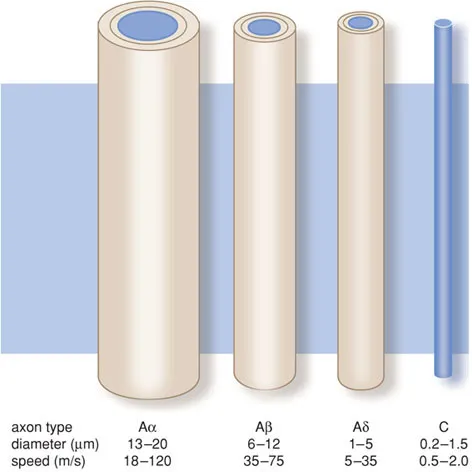
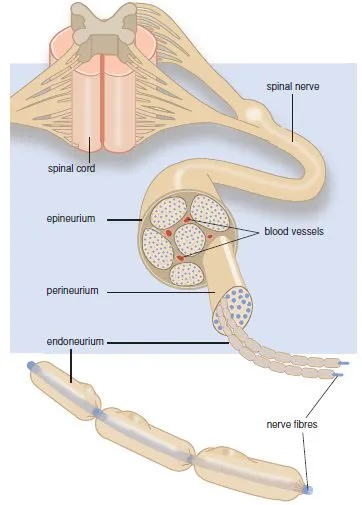
Nerves are bundles of nerve fibres held together by connective tissue (Figure 1.4). Each individual axon is surrounded by connective tissue (endoneurium). Bundles of nerve fibres form a fascicle, which is also held together by connective tissue (perineurium). A number of fascicles are held together again by connective tissue (epineurium), forming a nerve.
1.2.2 Impulse formation
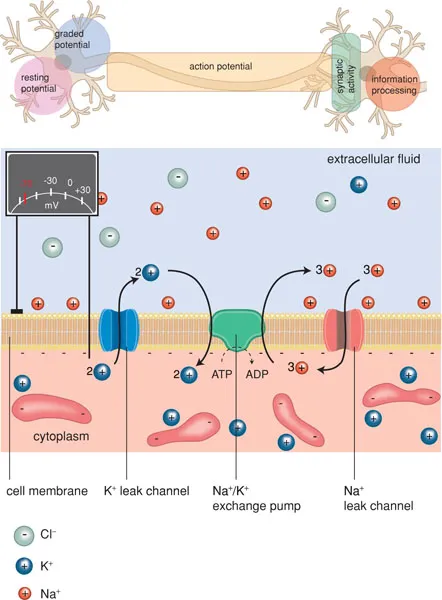
The generation and conduction of impulses in nerve fibres is a complicated process. In order to excite electrical impulses, a change in electrical charge must take place. Cells are surrounded by a semipermeable membrane that is only permeable to water. A selective ion pump actively pumps potassium ions into the cell and sodium ions out of the cell. This results in a concentration gradient of sodium and potassium ions over the membrane. The cell cytoplasm contains a high concentration of negatively charged proteins, which give the cell a negative charge compared with its environment. Extracellularly, negatively charged ions are also present, primarily chloride ions. On both sides of the membrane, the electrical charge is balanced by positively charged ions (sodium, potassium, calcium). Because the concentration of anions on the inside is slightly higher than on the outside, the number of cations inside will therefore be higher than outside. This causes a transmembrane potential difference of −60 mV, called the resting potential.
Figure 1.3 The nerve cell.
The membrane contains ion channels with an open and closed state (Figure 1.5). These channels can be activated by an electrical stimulus (‘voltage-gated’) or by a chemical stimulus (‘ligand-gated’) (see Box 1.2). When ion channels are open, ions move along the concentration gradient. At rest, primarily potassium channels are open, so that potassium ions try to leave the cell. However, the relative overload of anions in the cell (proteins) counteracts the outflow of cations. When the sodium channels of the membrane open, sodium ions will move in: in other words, the membrane has a hole.
The inflow of sodium ions distorts the electrical equilibrium, so that a local depolarisation occurs and potassium ions can leave the cell. This restores the balance between anions and cations (repolarisation). During depolarisation and the beginning of repolarisation, no new depolarisation can occur (refractory period).
Figure 1.4 The peripheral nerve.
When the local depolarisation is slight, the equilibrium is quickly restored (Figure 1.6). Only when the local depolarisation reaches a certain threshold value (approx. −50 mV), does an action potential appear. Thus there is an ‘all-or-none’ effect.
The height of the threshold value, necessary for an action potential to develop, is determined by several factors, such as the duration and strength of the depolarising stimulus and the status of the receptor. Through this, the voltage-gated sodium channels are opened, so that an influx of sodium occurs and the membrane polarity reverses.
Figure 1.5 Semi-permeable membrane with ion channels.
Box 1.2
Ion channels are of great importance for the generation, conduction and transfer of nerve impulses. Activation of these receptors may occur by an electrical stimulus (voltage-gated channels) or by a neurotransmitter (ligand-gated channels). Once activated the channel opens, which allows the passage of ions, causing a depolarisation of the cell membrane.
Voltage-gated ion channels are, amongst others, the fast sodium channels and calcium channels involved in impulse formation in the heart and in impulse conduction in the nerve fibres. Examples of ligand-gated ion channels are acetylcholine receptors, glutamate receptors and GABA receptors.
Figure B1.2 A–C
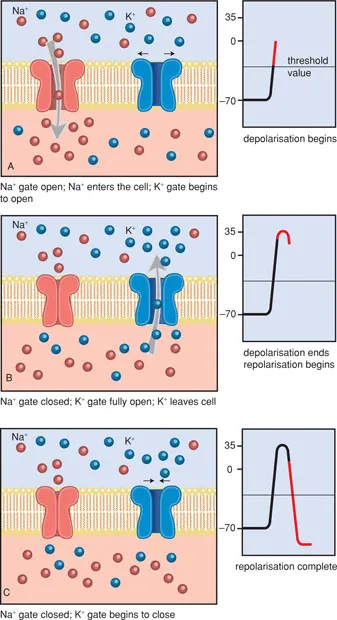
Figure B1.2 D Activation of a ligand-gated ion channel.
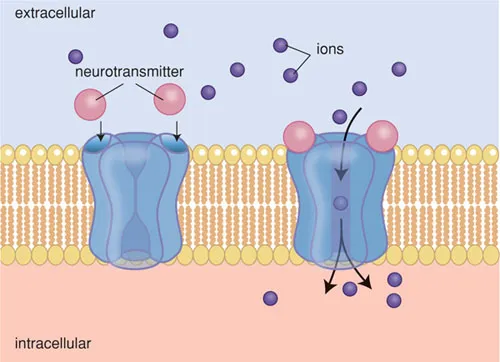
Figure 1.6 The action potential.
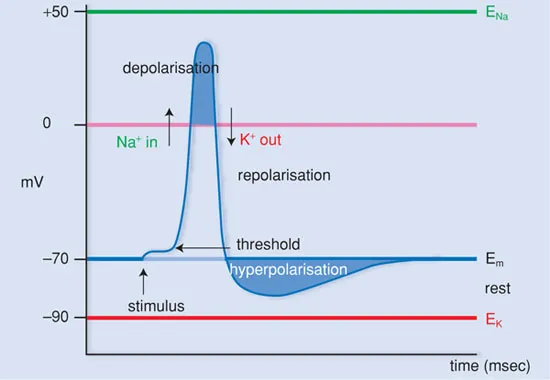
The sodium channels remain open only for approx. 1 millisecond, after which they close again. The p...