
- 1,160 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
This 2e of Toxoplasma gondii reflects the significant advances in the field in the last 5 years, including new information on the genomics, epigenomics and proteomics of T. gondii as well as a new understanding of the population biology and genetic diversity of this organism. T. gondii remains the best model system for studying the entire Apicomplexa group of protozoans, which includes Malaria, making this new edition essential for a broad group of researchers and scientists.
Toxoplasmosis is caused by a one-celled protozoan parasite known as T. gondii. The infection produces a wide range of clinical syndromes in humans, land and sea mammals, and various bird species. Most humans contract toxoplasmosis by eating contaminated, raw or undercooked meat (particularly pork), vegetables, or milk products; by coming into contact with the T. gondii eggs from cat feces; or by drinking contaminated water. The parasite damages the ocular and central nervous systems, causing behavioral and personality alterations as well as fatal necrotizing encephalitis. It is especially dangerous for the fetus of an infected pregnant woman and for individuals with compromised immune systems, such as HIV-infected patients.
- Completely updated, the 2e presents recent advances driven by new information on the genetics and genomics of the pathogen
- Provides the latest information concerning the epidemiology, diagnosis, treatment and prevention of toxoplasmosis
- Offers a single-source reference for a wide range of scientists and physicians working with this pathogen, including parasitologists, cell and molecular biologists, veterinarians, neuroscientists, physicians, and food scientists
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
The History and Life Cycle of Toxoplasma gondii
Jitender P. Dubey, Animal Parasitic Diseases Laboratory, Beltsville Agricultural Research Center, Agricultural Research Service, United States Department of Agriculture, Beltsville, Maryland, USA
Abstract
Toxoplasma gondii has a complex life cycle with multiple forms. Intermediate hosts such as humans are infected by sporozoites in oocysts or bradyzoites in pseudocysts whereas the sexual stages occur in the intestine of the definitive host, feline species. The parasite is unusual in that it does not need to pass through sexual stages or its definitive host for transmission to other species. This chapter summarizes the history of the discovery of the parasite, identification of the definitive host and transmission stages of the parasite, as well as other important discoveries in the basic biology and life cycle of T. gondii. This chapter also provides photomicrographs of the life cycle stages of this organism.
Keywords
tachyzoite; bradyzoite; sporozoite; sexual cycle; transmission; toxoplasmosis; diagnosis
Outline
1.1 Introduction
1.2 The Etiological Agent
1.3 Parasite Morphology and Life Cycle
1.3.1 Tachyzoites
1.3.2 Bradyzoite and Tissue Cysts
1.3.3 Enteroepithelial Asexual and Sexual Stages
1.4 Transmission
1.4.1 Congenital
1.4.2 Carnivorism
1.4.3 Faecal–Oral
1.5 Toxoplasmosis in Humans
1.5.1 Congenital Toxoplasmosis
1.5.2 Acquired Toxoplasmosis
1.5.2.1 Children
1.5.2.2 Toxoplasmosis in Adults
1.6 Toxoplasmosis in Other Animals
1.7 Diagnosis
1.7.1 Sabin–Feldman Dye Test
1.7.2 Detection of IgM Antibodies
1.7.3 Direct Agglutination Test
1.7.4 Detection of T. gondii DNA
1.8 Treatment
1.9 Prevention and Control
1.9.1 Serologic Screening During Pregnancy
1.9.2 Hygiene Measures
1.9.3 Animal Production Practices
1.9.4 Vaccination
Acknowledgements
References
Acknowledgements
I would like to thank Drs. Georges Desmonts (now deceased), David Ferguson, Jack Frenkel, H.R. Gamble, Garry Holland, Jeff Jones, and Jack Remington for their helpful discussions in the preparation of this manuscript.
1.1 Introduction
Infections by the protozoan parasite Toxoplasma gondii are widely prevalent in humans and other animals on all continents. There are many thousands of references to this parasite in the literature and it is not possible to give equal treatment to all authors and discoveries (Dubey, 2008). The objective of this chapter is, rather, to provide a history of the milestones in our acquisition of knowledge of the biology of this parasite (Fig. 1.1).
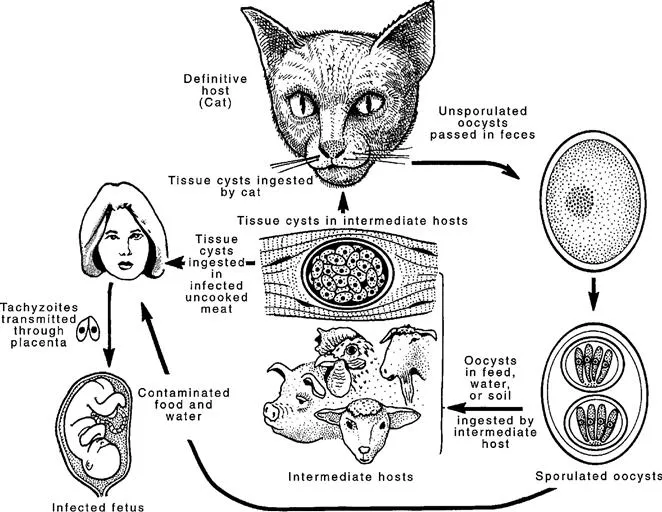
FIGURE 1.1 Life cycle of T. gondii.
1.2 The Etiological Agent
Nicolle and Manceaux (1908) found a protozoan in tissues of a hamster-like rodent, the gundi, Ctenodactylus gundi, which was being used for leishmaniasis research in the laboratory of Charles Nicolle at the Pasteur Institute in Tunis. They initially believed the parasite to be Leishmania, but soon realized that they had discovered a new organism and named it Toxoplasma gondii based on the morphology (Modern Latin Toxo = arc or bow, plasma = life) and the host (Nicolle and Manceaux, 1909). Thus, its complete designation is Toxoplasma gondii (Nicolle and Manceaux, 1908). In retrospect, the correct name for the parasite should have been Toxoplasma gundii; Nicolle and Manceaux (1908) had incorrectly identified the host as Ctenodactylus gondi. Splendore (1908, translated in 2009 into English) discovered the same parasite in a rabbit in Brazil, also erroneously identifying it as Leishmania, but he did not name it. It is a remarkable coincidence that this disease was first recognized in laboratory animals and was first thought to be Leishmania by both groups of investigators.
1.3 Parasite Morphology and Life Cycle
1.3.1 Tachyzoites
The tachyzoite (Frenkel, 1973) is lunate and is the stage that Nicolle and Manceaux (1909) found in the gundi (Fig. 1.2A). This stage has also been called trophozoite, the proliferative form, the feeding form, and endozoite. It can infect virtually any cell in the body. It divides by a specialized process called endodyogeny, first described by Goldman et al. (1958). Gustafson et al. (1954) first studied the ultrastructure of the tachyzoite. Sheffield and Melton (1968) provided a complete description of endodyogeny when they fully described its ultrastructure.
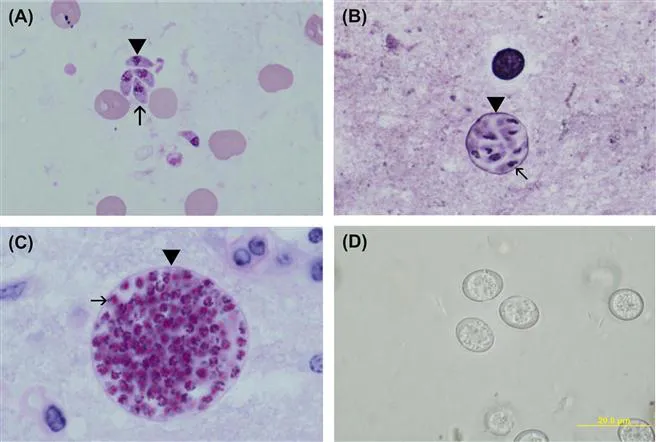
FIGURE 1.2 Life cycle stages of T. gondii.
A) Tachyzoites (arrowhead) in smear. Giemsa stain. Note nucleus dividing into two nuclei (arrow).
B) A small tissue cyst in smear stained with Giemsa and a silver stain. Note the silver-positive tissue cyst wall (arrow head) enclosing bradyzoites that have a terminal nucleus (arrow).
C) Tissue cyst in section, PAS. Note PAS-positive bradyzoites (arrow) enclosed in a thin PAS-negative cyst wall (arrowhead).
D) Unsporulated oocysts in cat faeces. Unstained.
A) Tachyzoites (arrowhead) in smear. Giemsa stain. Note nucleus dividing into two nuclei (arrow).
B) A small tissue cyst in smear stained with Giemsa and a silver stain. Note the silver-positive tissue cyst wall (arrow head) enclosing bradyzoites that have a terminal nucleus (arrow).
C) Tissue cyst in section, PAS. Note PAS-positive bradyzoites (arrow) enclosed in a thin PAS-negative cyst wall (arrowhead).
D) Unsporulated oocysts in cat faeces. Unstained.
1.3.2 Bradyzoite and Tissue Cysts
The term ‘bradyzoite’ (Greek brady = slow) was proposed by Frenkel (1973) to describe the stage encysted in tissues. Bradyzoites are also called cystozoites. Dubey and Beattie (1988) proposed that cysts should be called tissue cysts to avoid confusion with oocysts (Figs 1.2B,C). It is difficult to determine from the early literature who first identified the encysted stage of the parasite (Lainson, 1958). Levaditi et al. (1928) were apparently the first to report that T. gondii may persist in tissues for many months as ‘cysts’; however, considerable confusion between the term ‘pseudocysts’ (a group of tachyzoites) and tissue cysts existed for many years. Frenkel and Friedlander (1951) and Frenkel (1956) characterized cyst cytologically as containing organisms with a subterminal nucleus and periodic acid Schiff (PAS)-positive granules surrounded by an argyrophilic cyst wall (Fig. 1.2B,C). Wanko et al. (1962) first described the ultrastructure of the T. gondii cyst and its contents. Jacobs et al. (1960a) first provided a biological characterization of cysts when they found that the cyst wall was destroyed by pepsin or trypsin, but the cystic organisms were resistant to digestion by gastric juices (pepsin-HCl), whereas tachyzoites were destroyed immediately. Thus, tissue cysts were shown to be important in the life cycle of T. gondii because carnivorous hosts can become infected by ingesting infected meat. Jacobs et al. (1960b) used the pepsin digestion procedure to isolate viable T. gondii from tissues of chronically infected animals. When T. gondii oocysts were discovered in cat faeces in 1970, oocyst shedding was added to the biological description of the cyst (Dubey and Frenkel, 1976).
Dubey and Frenkel (1976) performed the first in-depth study of the development of tissue cysts and bradyzoites and described their ontogeny and morphology. They found that tissue cysts formed in mice as early as three days after their inoculation with tachyzoites. Cats shed oocysts with a short prepatent period (three to 10 days) after ingesting tissue cysts or bradyzoites, whereas after they ingested tachyzoites or oocysts the prepatent period was longer (≥18 days), irrespective of the number of organisms in the inocula (Dubey and Frenkel, 1976; Dubey, 1996, 2001, 2006) (Fig. 1.2D). Prepatent periods of 11–17 days are thought to result from the ingestion of transitional stages between tachyzoite and bradyzoite (Dubey, 2002, 2005).
Wanko et al. (1962), and Ferguson and Hutchison (1987) reported on the ultrastructural development of T. gondii tissue cysts. The biology of bradyzoites including morphology, development in cell culture in vivo, conversion of tachyzoites to bradyzoites, and vice versa, tissue cyst rupture, and distribution of tissue cysts in various hosts and tissues was reviewed critically by Dubey et al. (1998).
1.3.3 Enteroepithelial Asexual and Sexual Stages
Asexual and sexual stages were reported in the intestine of cats in 1970 (Frenkel, 1970) (Figs 1.3 and 1.4). Dubey and Frenkel (1972) described the asexual and sexual development of T. gondii in enterocytes of the cat and designated the asexual enteroepithelial stages as types A through E rather than as generations conventionally known as schizonts in other coccidian parasites (Figs 1.3 and 1.4). These stages were distinguished morphologically from tachyzoites and bradyzoites, which also occur in cat intestine (Fig. 1.3D). The challenge was to distinguish different stages in the cat intestine because there was profuse multiplication of T. gondii three days post-infection (Fig. 1.4A). The entire cycle was completed in 66 hours after feeding...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Dedication
- Preface to the First Edition
- Preface to the Second Edition
- List of Contributors
- Chapter 1. The History and Life Cycle of Toxoplasma gondii
- Chapter 2. The Ultrastructure of Toxoplasma gondii
- Chapter 3. Molecular Epidemiology and Population Structure of Toxoplasma gondii
- Chapter 4. Human Toxoplasma Infection
- Chapter 5. Ocular Disease due to Toxoplasma gondii
- Chapter 6. Toxoplasmosis in Wild and Domestic Animals
- Chapter 7. Toxoplasma Animal Models and Therapeutics
- Chapter 8. Biochemistry and Metabolism of Toxoplasma gondii: Carbohydrates, Lipids and Nucleotides
- Chapter 9. The Apicoplast and Mitochondrion of Toxoplasma gondii
- Chapter 10. Calcium Storage and Homeostasis in Toxoplasma gondii
- Chapter 11. The Toxoplasma gondii Parasitophorous Vacuole Membrane: A Multifunctional Organelle in the Infected Cell
- Chapter 12. Toxoplasma Secretory Proteins and Their Roles in Cell Invasion and Intracellular Survival
- Chapter 13. The Toxoplasma Cytoskeleton: Structures, Proteins and Processes
- Chapter 14. Interactions Between Toxoplasma Effectors and Host Immune Responses
- Chapter 15. Bradyzoite Development
- Chapter 16. Development and Application of Classical Genetics in Toxoplasma gondii
- Chapter 17. Genetic Manipulation of Toxoplasma gondii
- Chapter 18. Epigenetic and Genetic Factors that Regulate Gene Expression in Toxoplasma gondii
- Chapter 19. ToxoDB: An Integrated Functional Genomic Resource for Toxoplasma and Other Sarcocystidae
- Chapter 20. Comparative Aspects of Nucleotide and Amino Acid Metabolism in Toxoplasma gondii and Other Apicomplexa
- Chapter 21. Toxoplasma gondii Chemical Biology
- Chapter 22. Proteomics of Toxoplasma gondii
- Chapter 23. Cerebral Toxoplasmosis: Pathogenesis, Host Resistance and Behavioural Consequences
- Chapter 24. Innate Immunity to Toxoplasma gondii
- Chapter 25. Adaptive Immunity and Genetics of the Host Immune Response
- Chapter 26. Vaccination against Toxoplasmosis: Current Status and Future Prospects
- Epilogue
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Toxoplasma Gondii by Louis M. Weiss in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Infectious Diseases. We have over 1.5 million books available in our catalogue for you to explore.