
eBook - ePub
Hearing Loss
Causes, Prevention, and Treatment
Jos J. Eggermont
This is a test
Share book
- 426 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Hearing Loss
Causes, Prevention, and Treatment
Jos J. Eggermont
Book details
Book preview
Table of contents
Citations
About This Book
Hearing Loss: Causes, Prevention, and Treatment covers hearing loss, causes and prevention, treatments, and future directions in the field, also looking at the cognitive problems that can develop.
To avoid the "silent epidemic" of hearing loss, it is necessary to promote early screening, use hearing protection, and change public attitudes toward noise. Successful treatments of hearing loss deal with restoring hearing sensitivity via hearing aids, including cochlear, brainstem, or midbrain implants. Both the technical aspects and effects on the quality of life of these devices are discussed.
The integration of all aspects of hearing, hearing loss, prevention, and treatment make this a perfect one-volume course in audiology at the graduate student level. However, it is also a great reference for established audiologists, ear surgeons, neurologists, and pediatric and geriatric professionals.
- Presents an in-depth overview of hearing loss, causes and prevention, treatments, and future directions in the field
- Written for researchers and clinicians, such as auditory neuroscientists, audiologists, neurologists, speech pathologists, pediatricians, and geriatricians
- Presents the benefits and problems with hearing aids and cochlear implants
- Includes important quality of life issues
Frequently asked questions
How do I cancel my subscription?
Can/how do I download books?
At the moment all of our mobile-responsive ePub books are available to download via the app. Most of our PDFs are also available to download and we're working on making the final remaining ones downloadable now. Learn more here.
What is the difference between the pricing plans?
Both plans give you full access to the library and all of Perlego’s features. The only differences are the price and subscription period: With the annual plan you’ll save around 30% compared to 12 months on the monthly plan.
What is Perlego?
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 1000+ topics, we’ve got you covered! Learn more here.
Do you support text-to-speech?
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more here.
Is Hearing Loss an online PDF/ePUB?
Yes, you can access Hearing Loss by Jos J. Eggermont in PDF and/or ePUB format, as well as other popular books in Psicologia & Neuroscienza e neuropsicologia cognitiva. We have over one million books available in our catalogue for you to explore.
Information
Topic
PsicologiaPart I
The Basics
Outline
Chapter 1
Hearing Basics
Abstract
New findings about the structure and function of the auditory system continue to emerge. In this chapter, we describe, besides the “classical” textbook knowledge on the action of the cochlea and central nervous system, recent insights into the effects of subclinical noise exposures on the ribbon synapses, and insights into the protective effects of the efferent system for the effects of these subclinical noise exposures. This protective effect does not seem to include traumatic noise exposure, albeit that conditioning with moderate level sound that presumably includes activity of the efferent system induces such protection. We also pay close attention to new findings regarding the type II synapses innervating the outer hair cells. The growing knowledge about the efferent system in preventing or protecting against noise-induced and age-related hearing loss is extensively reviewed. The advent of powerful imaging techniques applied to the human auditory cortex has allowed a comparison with the detailed functional knowledge of these areas in nonhuman primates and suggests very strong similarities between them.
Keywords
Cochlear potentials; auditory nerve fibers; compound action potentials; ribbon synapses; parallel processing; efferent system; central auditory system; neural imaging
Hearing loss comprises reduced sensitivity for pure tones (the audiogram) and problems in the understanding of speech. The loss of sensitivity results from deficits in the transmission of sound via the middle ear and/or loss of transduction of mechanical vibrations into electrical nerve activity in the inner ear. Problems of speech understanding mainly result from deficits in the synchronization of auditory nerve fibers’ (ANFs) and central nervous system activity. This can be the result of problems in the auditory periphery but may also occur in the presence of nearly normal audiometric hearing. In order to appreciate the interaction of the audibility and “understanding” aspects of hearing loss, I will, besides presenting a condensed review of the auditory system, pay detailed attention to new findings pertaining to the important role of the ribbon synapses in the inner hair cells (IHCs), parallel processing in the ascending auditory system, and finally the importance of the efferent system.
1.1 Hearing Sensitivity in the Animal Kingdom
Every animal that grunts, croaks, whistles, sings, barks, meows, or speaks can hear. Most of the hearing animal species that we are familiar with are vertebrates, however insects also have keen hearing. For cicades and crickets that come as no surprise as these form choruses to enliven our nights. It may also turn out that the humble fruit fly, Drosophila, whose song is barely audible (Shorey, 1962), and the favorite of geneticists, may become important to elucidate the genetics of hearing loss (Christie et al., 2013).
A common way to quantify the hearing sensitivity (or loss) in humans is by way of the audiogram—a plot of the threshold level of hearing at a fixed series of (typically octave-spaced) frequencies between 125 Hz and 8 kHz when used in a clinical setting. In research settings, a wider and more finely spaced range of frequencies is employed (Fig. 1.1). In research settings, the just audible sound pressure level (dB SPL) is plotted, whereas in clinical settings the loss of sensitivity relative to a normalized value is represented (dB HL). To avoid confusion we call the research representation the “hearing field.”
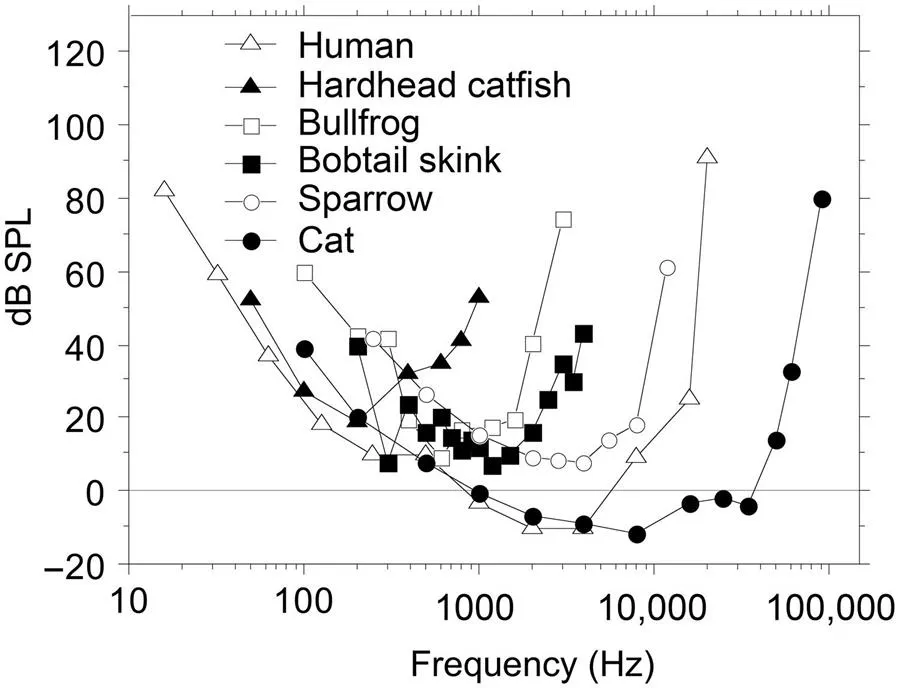
As Fig. 1.1 shows, hearing sensitivity differs considerably between vertebrates, even between mammals. Small mammals often have better high-frequency hearing than humans, with the 60 dB SPL upper limits of the hearing field ranging from 34.5 kHz for the Japanese macaque, and about 60 kHz for the cat to more than 120 kHz for the horseshoe bat (Heffner and Heffner, 2007). One reason for this variation may be that small mammals need to hear higher frequencies than larger mammals do in order to make use of sound localization cues provided by the frequency-dependent attenuating effect of the head and pinnae on sound. As a result, mammals with small heads generally have better high-frequency hearing than mammals with large heads, such as the elephant. Almost all mammals have poorer low-frequency hearing than humans, with the 60 dB lower limits ranging from 28 Hz for the Japanese macaque to 2.3 kHz for the domestic mouse (Heffner and Heffner, 2007; not shown). Only the Indian elephant, with a 60-dB low-frequency limit of 17 Hz, is known to have significantly better low-frequency hearing than humans, reaching into the infrasound range (Garstang, 2004).
Birds are among the most vocal vertebrates and have excellent hearing sensitivity. However, a striking feature of bird hearing is that the high-frequency limit, which falls between 6 and 12 kHz—even for small birds—is well below those of most mammals, including humans. Fig. 1.1 shows a typical bird audiogram represented by the sparrow. Among reptiles, lizards such as the bobtail skink (Fig. 1.1) are the best hearing species and are up to 30 dB more sensitive than alligators and crocodiles.
Anurans (frogs and toads) are very vocal amphibians: In specific parts of the year, depending on the species, their behavior is dominated by sound. As I wrote earlier (Eggermont, 1988): “Sound guides toads and frogs to breeding sites, sound is used to advertise the presence of males to other males by territorial calls, and sound makes the male position known to females through mating or advertisement calls. To have the desired effect these calls must be identified as well as localized. Frogs and toads are remarkably good in localizing conspecific males, especially when we take into account their small head size and the fact that hardly any of the territorial or mating calls has sufficient energy in the frequency region above 5 kHz to be audible at some distance.” Especially, the bullfrog’s threshold is relatively low at 10 dB SPL around 600 Hz (Fig. 1.1).
Teleost fishes, the largest group of living vertebrates, include both vocal and nonvocal species. This is especially evident for some by their intense sound production during the breeding season (Bass and McKibben, 2003). Except for the hardhead catfish (Fig. 1.1) that hears sounds at approximately 20 dB SPL for 200 Hz, most fishes have thresholds around 40 dB SPL, and with few exceptions do not hear sounds above 2 kHz (Popper and Fay, 2011).
Nearly all insects have high-frequency hearing (Fonseca et al., 2000). For instance, the hearing ranges for crickets are 0.1–60 kHz, for grasshoppers 0.2–50 kHz, for flies 1–40 kHz, and for cicades 0.1–25 kHz. Tiger moths are typically most sensitive to ultrasound frequencies between 30 and 50 kHz. The frequency sensitivity of the ears of moth species is often matched to the sonar emitted by the bats preying upon them (Conner and Corcoran, 2012).
1.2 The Mammalian Middle Ear
“The auditory periphery of mammals is one of the most remarkable examples of a biomechanical system. It is highly evolved, with tremendous mechanical complexity” (Puria and Steele, 2008).
Transmission of sound energy from air to fluid typically results in considerable loss as a result of reflection from the fluid surface and estimated at about 99.7% of the incoming energy. This is compensated by the pressure gain provided by the ratio of the areas of the tympanic membrane (TM) (typical 0.55 cm2) and the stapes footplate (typical 0.032 cm2) for human, which is approximately 17, and the lever action of the middle ear bones which contributes a factor approximately 1.3 (Dallos, 1973). This would theoretically result in a combined gain of a factor 22 (about 27 dB). In practice, the gain is considerably less and maximal between 20 and 25 dB in the 800–1500 Hz range (Dallos, 1973).
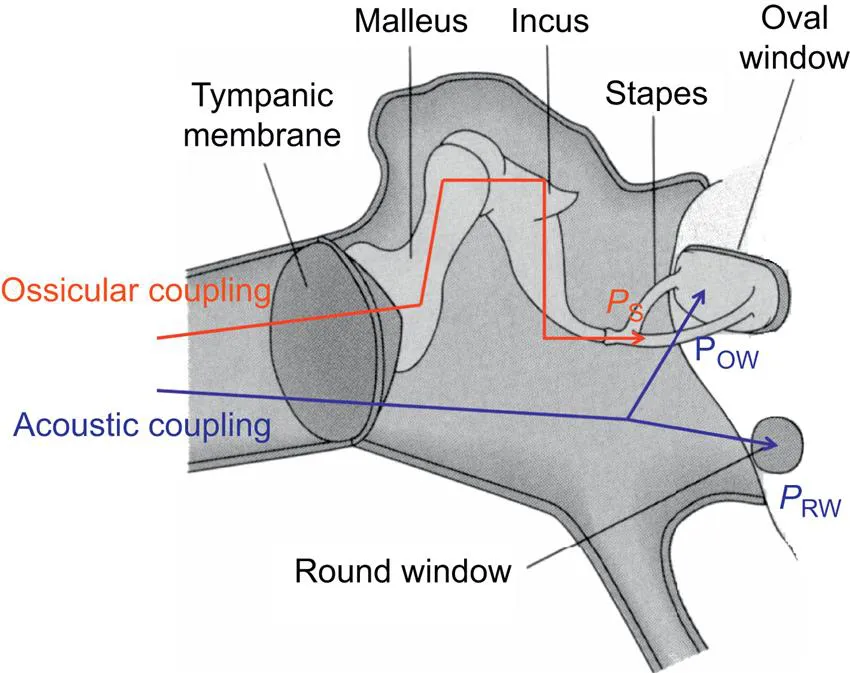
Merchant et al. (1997) extensively described the middle ear action as the result of two mechanisms: ossicular and acoustic coupling. Ossicular coupling incorporates the gain in sound pressure that occurs through the TM and ossicular chain. In the normal middle ear, sound pressure in the ear canal results in TM motion transmitted through the malleus and incus to produce a force at the stapes head (Fig. 1.2). This force applied over the area of the footplate produces a pressure PS. PS represents the ossicular coupling of sound to the cochlea. Acoustic coupling refers to the difference in sound pressures acting directly on the oval window (OW), POW, and round window (RW), PRW. In normal ears, acoustic coupling, ΔP=(POW−PRW), is negligibly small, but it can play a significant role in some diseased middle ears (Peake et al., 1992).
1.3 The Mammalian Inner Ear
Until 1971, the basilar membrane (BM) was considered to be a linear device with broad mechanical tuning, as originally found already in the 1940s by von Békésy (1960). The bridge to the narrow ANF tuning was even long thereafter considered the result of a “second filter” (Evans and Klinke, 1982). Thus, it took a while before the results from Rhode (1971) indicating that the BM was a sharply tuned nonlinear filtering device were accepted. Appreciating these dramatic changes in viewing the working of the cochlea, Davis (1983) wrote: “We are in the midst of a major breakthrough in auditory physiology. Recent experiments force us, I believe, to accept a revolutionary new hypothesis concerning the action of the cochlea namely, that an active process increases the vibration of the basilar membrane by energy provided somehow in the organ of Corti.” Then, another crucial discovery was that the outer hair cells (OHCs), in response to depolarization, were capable of producing a m...