Physiology of Human Reproduction provides students with a concise and accessible overview of more than 200 vital concepts, from the basic physiology of the male and the nonpregnant female, to fertilization, embryonic and fetal growth, labor, lactation, and more. Presented in a readable style, key terms are highlighted throughout the main text to enable students to quickly find a concept and read the appropriate information.
Whether reading the book from cover to cover, or using a focused approach to learn about specific concepts, readers will find this textbook to be an invaluable tool for increasing their understanding of human reproduction. An essential companion for standard Anatomy and Physiology courses, this student-friendly textbook:
Covers physiology of the male, the physiology of the nonpregnant female, pregnancy and lactation, and age-related changes such as menopause
Discusses pregnancy, birth control, and the reproductive system in childhood, adolescence, and puberty
Describes the anatomy, physiology, and phases of the human sexual response
Explains genetic conditions and disorders including androgen insensitivity syndrome and Kallman's syndrome
Physiology of Human Reproduction is a must-have learning guide for students in the medical and life sciences, including medicine, nursing, biology, physiology, and biomedicine, as well as those in courses covering human reproduction and pregnancy.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
[See Appendix for a review of male reproductive anatomy.]
1.1 Three Basic Requirements for Fertility in the Male
For a man to be fertile, he must fulfill three basic requirements [1]: produce semen containing sufficient numbers of healthy sperm, achieve an erection of sufficient rigidity to enter the vagina during intercourse, and be capable of ejaculating in a way that deposits semen within the vaginal canal.
Unlike the complex cyclicity of the female, male sexual function is fairly invariant. As in the female, the male reproductive system is regulated and maintained by the endocrine system, specifically the hypothalamic–pituitary–gonadal axis. The key hormones involved include: hypothalamic gonadotropin‐releasing hormone (GnRH), two gonadotropins (LH, FSH) secreted from the anterior pituitary (also called the adenohypophysis), and two androgenic sex steroids (testosterone – T, and its metabolite dihydrotestosterone, or DHT).
1.2 Endocrinology of the Male Reproductive System
The reproductive system in both sexes is driven by the pulsatile release of GnRH from the hypothalamus. GnRH [2] has a short (less than four minutes) half‐life and consists of 10 amino acids that are cleaved off of a larger molecule (pre‐proGnRH) whose production is coded for by chromosome 8. Once released, the GnRH decapeptide quickly travels the few millimeters to the anterior pituitary (adenohypophysis) via a specialized capillary portal system, where it binds to pituitary cells called gonadotropes (or gonadotrophs) and stimulates the release of luteinizing hormone (LH) and follicle‐stimulating hormone (FSH) [3] – two gonadotropins named for their actions in the female. Like GnRH, the gonadotropins are also released in pulses and, as the term trophic suggests, their function is to “nourish” (Gr), i.e. to maintain the structure and function of the testes and ovaries.
Although pulsatile release [4] of GnRH, and of LH and FSH occurs in both the male and female, the patterns of pulsatility differ, and are used by the body to encode their physiological actions. Like a combination of AM and FM radios, their amplitude (how much hormone is released in each pulse) and frequency (time between pulses) vary over time and changes in these parameters allow the body to orchestrate the complexity of the menstrual cycle in nonpregnant women, and to maintain testosterone secretion and reproductive function in the adult male.
Although LH and FSH are structurally different, both are glycoproteins (chains of amino acids linked to sugars). Because these complex and three‐dimensional molecules are charged (polar), they are unable to cross the cell membrane and rely instead on cellular recognition via receptors expressed on the plasma membrane.
Gonadotropin binding to the cell membrane is the first messenger [5]. This event activates the receptor and turns on ionic and/or enzymatic pathways inside the cell to generate the second messengers (e.g. calcium and kinases) that regulate its various functions such as secretion, metabolism, and hormone production.
Leydig cells [6] interspersed between the seminiferous tubules (also called interstitial cells for this reason) are the primary target of LH. The hormone and the cell type having a first letter in common (LH‐Leydig) is an easy way to remember this specificity, and the main action of LH in males [7] is to stimulate the Leydig cells to produce testosterone [8]. Once released, testosterone diffuses into a neighboring seminiferous tubule, within which sperm production occurs, and initiates paracrine actions on the Sertoli cells [9] that form the inner lining of each seminiferous tubule to assist in the highly ordered process of spermatogenesis that results in the creation of 100–200 million new sperm daily.
Alternatively, testosterone released from the Leydig cell can be absorbed into the bloodstream via one of the many capillaries that course between the seminiferous tubules and among the Leydig cells. Once testosterone enters the vascular compartment, it travels throughout the body to exert endocrine actions on many tissues such as skeletal muscle or the brain.
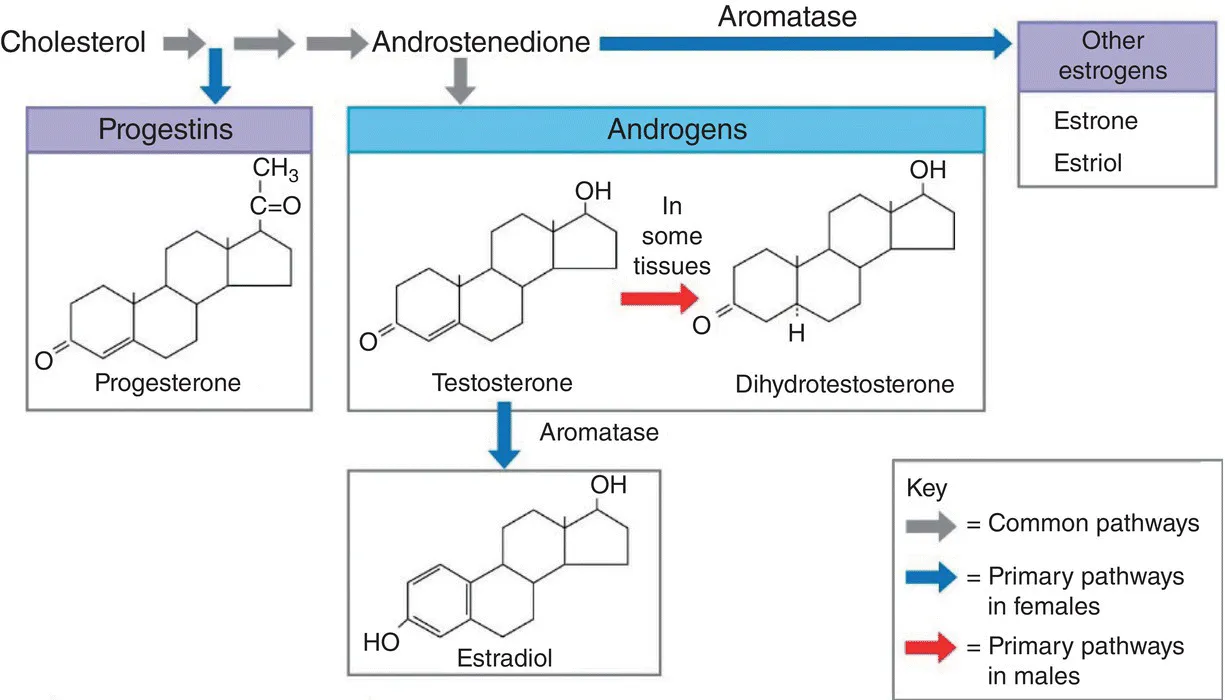
While there are other androgenic variants and metabolites of this steroid (the most common being DHT), all androgens have 19 carbons. In contrast, progesterone has 21, and estrogen 18. The three main types of sex steroids [10] – progestogens, androgens, and estrogens – are classified by their number of carbons. In addition, there are several steroidal enzymatic pathways by which one steroid may be converted into another. For example, testosterone can be produced from progesterone via 17α‐hydroxylation followed by cleavage of two carbons, and estrogen can be made from testosterone via the process of aromatization [11], which involves the appropriately named enzyme aromatase (Figure 1.1).
Interestingly, aromatization occurs in some of the neurons of the male brain, so that some of the central actions of testosterone – the male sex hormone – are actually carried out via its conversion into estrogen [12], the female sex hormone. Being small and highly lipid‐soluble molecules, all steroids diffuse into their target cells and enter the nucleus, where they combine with a nuclear receptor and activate or repress gene transcription. As we learn more about them, however, it is clear that there are also receptors in the cell membrane and, in the case of androgens, these are called membrane androgen receptors, or mARs. The activation of these G‐protein‐coupled receptors (GPCRs) leads to the generation of second messengers much like that elicited by the binding of the gonadotropins and all other peptide hormones. Acting in concert with promoter elements, testosterone modulates gene transcription to facilitate spermatogenesis and stimulates a range of Sertoli cell metabolic and secretory activities.
Figure 1.1 Steroidal biochemical pathways.
Once it reaches the testes, FSH [13], the other pituitary gonadotropin, diffuses from capillary across the connective tissue wall of the seminiferous tubule where it binds to the Sertoli cell membrane and elicits a cascade of second messengers. In response to FSH, Sertoli cells also make a peptide hormone of their own, called inhibin [14], which diffuses out of the tubule, enters the bloodstream and regulates FSH secretion through a negative feedback mechanism (just like testosterone regulates LH).
FSH and testosterone thereby regulate numerous Sertoli cell functions [15] in a cooperative manner. For example, Sertoli cells nurture the developing sperm by secreting nutrients (which is why they are often also called “nurture cells”). They also secrete a fluid into the lumen of the tubule that helps wash the sperm out of the tubules into the epididymis, and produce and secrete a protein called androgen‐binding protein, or ABP, into the lumen of the tubule. By continually binding and releasing many testosterone molecules, ABP acts like a testosterone “sponge” and sets up a dynamic equilibrium that allows testosterone concentrations within the tubule to be as much as 50 times higher than those in the systemic circulation.
Sertoli cells also phagocytize the cellular debris shed during spermiogenesis, which we describe later, and produce immunoregulatory molecules that suppress the immune response within the seminiferous tubules. This is important, since the immune system would perceive sperm as being antigenic and mount an autoimmune response by triggering autoantibody production.
A second Sertoli‐based anti‐immune mechanism is physical: each individual Sertoli cell is connected to its neighbors via tight junctions, which act to “zipper” together the cells into a sheet‐like intratubular barrier called the blood–testis barrier [16], preventing the passage of immune and other cells into the immunologically privileged intratubular compartment.
One final consideration related to the spatial relationships between the reproductive cells of the male and the female [17] may be helpful in remembering which hormones act on which cells. We have already considered the tripartite reproductive “unit” of the male as consisting of (i) Leydig cells outside, and (ii) Sertoli cells and (iii) sperm‐forming cells inside the seminiferous tubule.
In females, the analogous reproductive units are the mature ovarian follicles. The outer follicular shell (analogous to the Leydig cells in the male) is composed of theca cells while the inner one (analogous to Sertoli cells) consists of granulosa cells. Like Leydig cells, theca cells bind LH and produce testosterone. On the other hand, the inner granulosa cells that are closest to the egg (like the Sertoli cells closest to the sperm), bind FSH and take up the testosterone released from the neighboring outer theca cells.
Like Sertoli cells, granulosa cells also secrete the hormone inhibin, which enters the circulation and provides a negative feedback on pituitary FSH release. The one important difference is what happens to the testosterone that enters a granulosa cell: because granulosa cells have an abundance of aromatase, testosterone (19C) is converted to estrogen (18C) and released to diffuse out of the follicle and into the bloodstream where, and, in addition to exerting many its many endocrine effects on the uterus, brain, breasts, and other organs, the estrogen provides a negative feedback on pituitary LH release. To round out the analogy, note that Sertoli cells also contain some aromatase, and some of the Leydig cell‐derived testosterone is converted into 17‐beta estradiol to also help direct spermatogenesis.
1.3 Physiological Actions of Testosterone and Related Androgens
The testosterone that passes into the capillaries of the testis and circulates throughout the body has multiple and diverse ...
Table of contents
Cover
Table of Contents
Title Page
Copyright Page
Acknowledgement
Key Concepts and Terms
Introduction
1 The Adult Male
2 The Nonpregnant Adult Female
3 Pregnancy,Parturition,Lactation
4 The Human Sexual Response
5 Age‐Related Changes in theReproductive System
Appendix A: Detailed Anatomy of the Male and Female Reproductive Systems
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Physiology of Human Reproduction by George Osol in PDF and/or ePUB format, as well as other popular books in Medicine & Physiology. We have over 1.5 million books available in our catalogue for you to explore.