The Geometric Induction of Bone Formation describes new biomimetic biomaterials that offer mechanistic osteogenic surfaces for the autonomous and spontaneous induction of bone formation without the addition of osteogenic soluble molecular signals of the transforming growth factor-? supergene family. The chapters frame our understanding of regenerative medicine in primate species, including humans. The goal is to unravel the fundamental biological mechanisms of bone formation unique to non-human and human primates. The broad target audience dovetails with several disciplines both in the academic and private biotech sectors primarily involved in molecular biology, tissue biology, tissue engineering, biomaterial science, and reconstructive, orthopedic, plastic, and dental surgery.
Key Features
Includes outstanding images of undecalcified whole mounted sections
Summarizes non-human primate research – ideal for clinical translation
Reviews methods for creating devices capable of making bone autonomously, i.e. an intrinsically osteo-inductive bioreactor and/or biomaterial
Describes the spontaneous induction of bone formation including a whole spectrum of tissue biology, from basic molecular biology to clear-cut morphology and pre-clinical application in non-human primate species
Intended for audiences in both academic research and the biotech industry
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Functionalized Biomimetic Surfaces beyond Morphogens and Stem Cells
Ugo Ripamonti and Laura C. Roden
1.1 Tissue Induction and Morphogenesis
Serendipitously, one of the authors came across key papers on the induction of bone formation, the paper in the Proceedings of the National Academy of Sciences USA by Reddi and Huggins (1972) and the paper of Urist in Science (1965). Further searches led to two critical papers by Sampath and Reddi in the Proceedings of the National Academy of Sciences USA 1981 and 1983. Both papers described the induction of bone formation as a combinatorial molecular protocol recombining, or reconstituting, soluble and insoluble signals to trigger the ripple-like cascade of the induction of bone formation (Sampath and Reddi 1981; Sampath and Reddi 1983). The critical importance of the dissociative extraction and reconstitution of the extracellular matrix of bone is what propelled the experimental and clinical progression of the “bone induction principle” (Urist et al. 1967) from pre-clinical to clinical studies (Sampath and Reddi 1981; Sampath and Reddi 1983; Ripamonti and Reddi 1995; Reddi 1997; Reddi 2000; Ripamonti et al. 2001; Ripamonti 2006; Ripamonti et al. 2006; Ripamonti et al. 2007).
However, it was the paper by Piecuch entitled “Extraskeletal implantation of a porous hydroxyapatite ceramic” published in the Journal of Dental Research (1982) that inspired the author’s study of the biology of the incorporation of such macroporous biomatrices not only in bony sites but also in intramuscular heterotopic sites of the Chacma baboon Papio ursinus. Macroporous hydroxyapatite constructs derived from corals, with an average pore size of 500 µm, were prepared by Interpore International (Irvine, CA) and implanted in calvarial defects and in rectus abdominis intramuscular sites (Ripamonti 1991; Ripamonti et al. 2001; Ripamonti 2009). Experiments with these constructs demonstrated the spontaneous osteoinductivity of the macroporous constructs (Ripamonti 2006; Ripamonti 2009; Ripamonti 2017).
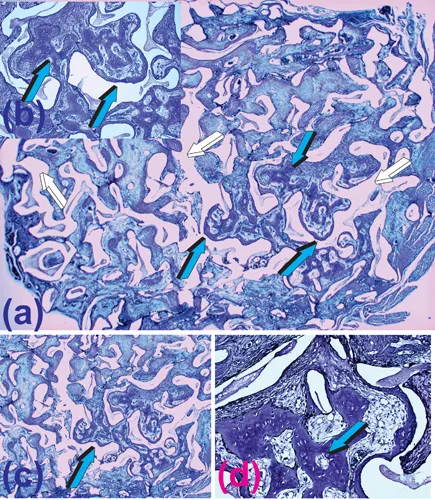
When implanted in heterotopic intramuscular sites, the coral-derived constructs initiated the induction of bone formation within the macroporous spaces. The induced bone was evident within the pores of the specimens’ harvested 90 days post-intramuscular implantation (Fig. 1.1) (Ripamonti 1990; Ripamonti 1991). Bone had formed tightly attached to the macroporous surfaces and extended with fine trabeculae across the macroporous spaces, supported by a closely associated rich vascular network (Fig. 1.1). These coral-derived substrata were later described as “macroporous bioreactors” (Ripamonti et al. 2009; Ripamonti et al. 2010; Ripamonti 2017), but it took almost 25 years to resolve the morphological data to understand and unravel the mechanism of the spontaneous induction of bone formation within the macroporous constructs (Klar et al. 2013).
FIGURE 1.1 Bone induction and morphogenesis spontaneously initiating within the macroporous spaces of a coral-derived bioreactor implanted heterotopically in the rectus abdominis muscle of a Chacma baboon Papio ursinus on day 90 after intramuscular implantation. (a) The spontaneous induction of bone formation without the exogenous application of osteogenic proteins of the transforming growth factor-β (TGF-β) supergene family. White arrows indicate the calcium phosphate biomatrix dissolved after histological processing (Ripamonti 1991). Vascularized trabeculae of newly formed woven bone (light blue arrows) formed by day 90 within the macroporous spaces supported by prominent angiogenesis (inset b). (c, d) High-power views of (a) detailing the induction of hyper cellular woven bone formation (light blue arrows) tightly attached to the biomimetic matrix substratum. Decalcified sections cut at 6 µm from paraffin-embedded blocks stained with toluidine blue in 70% ethanol. The images are from histological section 435/88 in honour of Barbara van den Heever who prepared impeccable sections after several experiments in Papio ursinus and alerted the writer to the morphogenesis of bone within the macroporous spaces of that section (Ripamonti 1991).
In collaboration with the Council for Scientific and Industrial Research (CSIR), Pretoria, we designed and tested highly crystalline sintered hydroxyapatites that also resulted in the spontaneous induction of bone formation (Ripamonti et al. 1999). This was par force the scientific evolution to further understand the induction of bone formation in coral-derived macroporous bioreactors (Ripamonti 1990; Ripamonti 1991; Ripamonti 1993; Ripamonti et al. 1993; Ripamonti 1996) and highly sintered crystalline hydroxyapatites for potential clinical applications (Ripamonti 1994; Ripamonti et al. 1999; Ripamonti 2004; Ripamonti et al. 1995; Ripamonti et al. 1997; Ripamonti et al. 2000; Ripamonti and Kirkbride 2001).
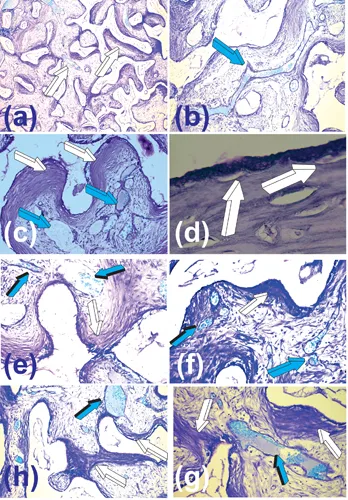
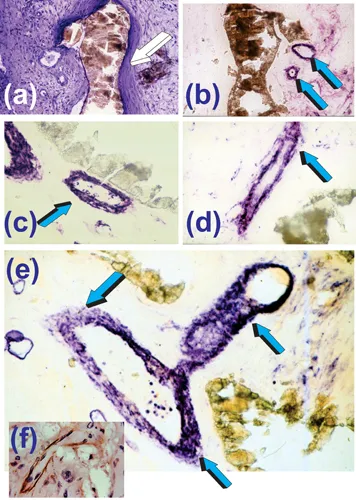
Experiments described in Ripamonti (1991) and Ripamonti et al. (1993) revealed the critical role of mesenchymal collagenous condensations that developed by day 30 post-implantation (Fig. 1.2). This was followed by detectable alkaline phosphatase expression by day 60 at the hydroxyapatite interface. These results indicated morphogenetic events pre-dating the induction of bone formation (Fig. 1.2) (Ripamonti 1990; Ripamonti 1991; Ripamonti et al. 1993). Furthermore, undecalcified sections showed alkaline phosphatase staining of the invading vasculature in close association with the coral-derived substrate by day 30 (Fig. 1.3). The stained multicellular layers of the invading capillaries were identified as the “osteogenetic vessels” described by Trueta (Trueta 1963). Laminin, a prominent vascular basement membrane component significantly associated with differentiation of the osteogenic phenotype (Foidart and Reddi 1980; Wlodarski and Reddi 1986), was also localized within the invading capillaries on days 30 and 60 (Fig. 1.3f).
FIGURE 1.2 Self-inducing macroporous biomimetic matrices initiating tissue induction and morphogenesis within the macroporous spaces on day 30 after heterotopic implantation, leading to the spontaneous and/or intrinsic induction of bone formation (Ripamonti 1990; Ripamonti 1991; Ripamonti et al. 1993; Ripamonti 1996). (a) Low-power view of a macroporous bioreactor harvested on day 30 after rectus abdominis implantation with differentiation of mesenchymal collagenous condensations at the hydroxyapatite interface (white arrows). (b) High-power view detailing the prominent vascular invasion (light blue arrow) within the fibrovascular tissue invading the macroporous spaces of the coral-derived bioreactor. (c) Collagenous condensations tightly attached to the macroporous substratum (white arrows) supported by prominent vascular invasion and angiogenesis (light blue arrow). (d) Early differentiating events at the hydroxyapatite interface with cellular differentiation towards the osteogenic phenotype (white arrows) in tight relationship with the substratum on day 30 after heterotopic implantation. (e,f,g,h) Patterns of vascular invasion, collagenous condensations’ distribution and alignment (white arrows) within the macroporous spaces of the coral-derived biomimetic matrices. There is always prominent angiogenesis (light blue arrows) continuously supporting tissue induction and morphogenesis within the macroporous spaces. Decalcified sections cut at 6 µm from paraffin-embedded blocks harvested and processed on day 30 after heterotopic implantation and stained with toluidine blue in 70% ethanol.
FIGURE 1.3 Early morphogenetic events on day 30 leading to the induction of bone formation by macroporous coral-derived bioreactors implanted in the rectus abdominis muscle of the Chacma baboon Papio ursinus. (a) Mesenchymal condensations on day 30 after heterotopic implantation (white arrow). (b) Consecutive serial section approximately 20 µm from (a) showing alkaline phosphatase staining (light blue arrows) of the invading osteogenetic vessels of Trueta’s definition (Trueta 1963). There is, however, a lack of alkaline phosphatase staining of the collagenous condensations against the hydroxyapatite interface. (c,d,e) Significant alkaline phosphatase staining (light blue arrows) of the invading sprouting capillaries within the macroporous spaces in tight relationship (c) with the macroporous biomimetic matrix. Note in (e) several layers of endothelial and perivascular cells expressing alkaline phosphatase whilst invading the macroporous spaces. Inset (f) immunolocalization of laminin within the basement membrane of an invading capillary (Ripamonti et al. 1993). Undecalcified sections from tissue blocks fixed in 70% ethanol, embedded, undecalcified, in historesin (LKB Bromma, Sweden), and cut at 3–5 µm (Ripamonti et al. 1993).
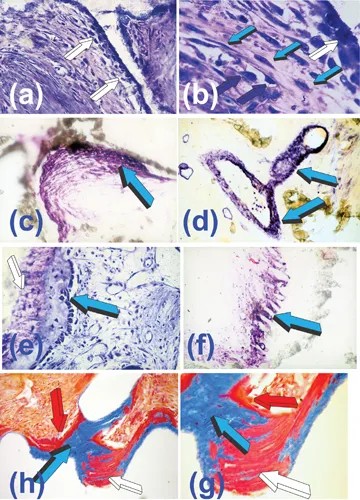
Inspired by Trueta’s paper on the role of the vessels in angiogenesis (Trueta 1963), we hypothesized that the alkaline phosphatase expression within the endothelial and sub-endothelial compartments of the osteogenetic vessels might provide a temporally regulated flow of bone precursor cells (Ripamonti et al. 1993). These precursor cells would be capable of the expression of the osteogenic phenotype when in contact with the calcium phosphate-based matrix of the implanted bioreactors (Figs. 1.3, 1.4). The morphological and biochemical data from these implants support this hypothesis (Figs. 1.4a,b) (Ripamonti et al. 1993; Ripamonti 2009). The induction and alignment of mesenchymal tissue condensations against the calcium phosphate-based surfaces are critical for the subsequent induction of bone formation (Ripamonti 1990; Ripamonti 1991; Ripamonti et al. 1993) (Figs. 1.2, 1.4).
FIGURE 1.4 Angiogenesis, capillary sprouting, cell migration and differentiation from the angiogenetic microenvironment to the bone induction microenvironment initiated by cellular differentiating and de-differentiating events by the “morphogenetic” and “osteogenetic vessels” of Aristotle’s (Crivellato et al. 2007) and Trueta’s definitions (Trueta 1963) at the osteogenetic microenvironment (white arrow in b). Vascular invasion and capillary sprouting provide the molecular, cellular and morphological templates for cell migration and differentiation with the induction of bone formation against the hydroxyapatite interface. (a,b) High-power views of the angiogenetic and osteogenetic microenvironments initiated by the calcium phosphate-based surface topography of the heterotopically implanted macroporous bioreactors on day 60 after heterotopic implantation. Middle-power view (a) with osteoblastic-like cell differentiation (white arrows) at the hydroxyapatite interface. (b) Detail of (a) highlighting osteoblastic-like cells differentiation at the interface (white arrow) and cellular trafficking in the angiogenetic compartment with highly hyperchromatic endothelial nuclei (dark blue arrows) seemingly migrating and de-differentiating from the penetrating vessels (light blue arrows) into the osteogenic microenvironment, providing a continuous flow of cellular progenitors for the...
Table of contents
Cover
Half-Title
Title
Copyright
Dedication
Contents
Acknowledgements
Author
List of Contributors
Introduction
Chapter 1 The New Frontiers in Bone Tissue Engineering: Functionalized Biomimetic Surfaces beyond Morphogens and Stem Cells
Chapter 2 The Induction of Bone Formation: When and Why Bone Forms and Sometimes Repairs and Regenerates: The Enigmatic Myth of Bone Tissue Engineering and the Dream of Regenerative Medicine
Chapter 3 The Induction of Bone Formation and the Osteogenic Proteins of the Transforming Growth Factor-β Supergene Family: Pleiotropism and Redundancy
Chapter 4 Coral-Derived Hydroxyapatite-Based Macroporous Bioreactors Initiate the Spontaneous Induction of Bone Formation in Heterotopic Extraskeletal Sites: Morphological Time Studies
Chapter 5 Concavities of Crystalline Sintered Hydroxyapatite-Based Macroporous Bioreactors Initiate the Spontaneous Induction of Bone Formation
Chapter 6 Molecular Pathways Regulating the Geometric Induction of Bone Formation: Synthetizing and Embedding Osteogenic Proteins into Nanotopographic Geometries
Chapter 7 The Spontaneous Induction of Bone Formation by Intrinsically Osteoinductive Bioreactors for Human Patients: Osteoinductive Hydroxyapatite-Coated Titanium Implants
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Geometric Induction of Bone Formation by Ugo Ripamonti in PDF and/or ePUB format, as well as other popular books in Scienze biologiche & Anatomia. We have over 1.5 million books available in our catalogue for you to explore.