![]()
Chapter 1
The Human Visual System
The visual systems of humans and other vertebrates are so complex and ingenious in the way that they operate that they were offered as a reason for doubting the theory of evolution given in 1859 by Charles Darwin (1809–1882). The argument put forward was that it was inconceivable that such a complicated system could arise in a gradual incremental way via mutations and natural selection. The visual system has several components and all of those components have to be present for the system to operate effectively. The opponents of Darwin argued that the human visual system can only have been a consequence of intelligent design and Darwin accepted that it was a case that needed to be answered — although he was adamant that evolutionary processes were capable of doing what was needed. In this chapter we shall begin with a description of the human visual system and then, in the following chapter, consider by what processes it could have evolved from a simple origin.
1.1. The Optical System
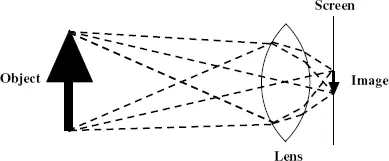
The human visual system has three distinct components: an optical system that produces an image; a neural network system that converts that image into electrical impulses; and an interpretation system, located in the brain, which converts the electrical signals into the sensation by which the object being imaged is visualized. The optical system is very simple and produces an image in the same way as is done by a simple convex lens. This is illustrated in Fig. 1.1 where the image of an arrow is projected onto a screen.
Figure 1.1 Producing an image on a screen with a simple convex lens.
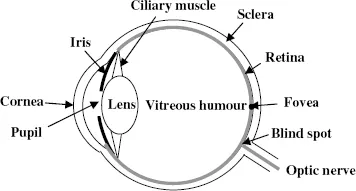
Figure 1.2 A schematic representation of a human eye.
The lens operates in such a way that rays of light, leaving a point on the object, travelling in different directions but all passing through the lens, are brought together at a point on the screen. From this diagram we can understand the conditions for producing a sharp image. If the screen were moved either forwards or backwards from the position shown then the light from a point on the object would be spread out into a circular patch and the image would be fuzzy. The same effect would come about as the result of keeping the position of the lens and screen constant but using a lens of a different focal length that bent the light differently.
Figure 1.2 shows a very simplified diagram of the structure of the human eye, and we now relate its operation in producing a sharp image to that of the simple lens. The combination of the cornea and lens produces an image on the retina, which is the equivalent of the screen in Fig. 1.1. However, for the eye, the distance of objects for which sharp images must be produced is variable, from very close when reading a book to very far when viewing a distant landscape. In the case of a lens and screen, if the distance of the object is changed then the distance of the screen from the lens can be changed to produce a sharp image. For the eye, the lens-to-screen distance is fixed so the only way to produce a sharp image is to change the focal length of the lens. This is done by means of the ciliary muscles that, by squeezing the lens, can change its shape and hence its focal length. The contribution of the cornea in the focusing process does not change but in a normal eye the total possible variation in the focal length of the lens is sufficient to allow clear images of both near and far objects to be produced. This variation is called accommodation and for some individuals it is unable to cover the full range of distances. Those who can see only close objects in focus suffer from short-sightedness, or myopia, while those able to see only far objects in focus suffer from far-sightedness, or hyperopia. In both cases spectacles can usually deal with the problem. Another common problem is that of astigmatism in which the curvature of the cornea is different in different directions. This means that light rays coming from the object and lying in different planes cannot all be brought into focus simultaneously. It is a common fault but normally so slight that it causes no difficulties to an individual. Where it is more severe then, once again, suitable spectacles can compensate for it.
The bulk of the eye is filled with a transparent gel-like material known as the vitreous humour. The sclera, seen in Fig. 1.2, together with the cornea, forms its outer container. While the cornea is transparent, as it must be to transmit light, the sclera is a milky-white colour — indeed, it is what is commonly called the white of the eye. The eye functions most efficiently at moderate light levels and the iris, the coloured disk that gives eye colour, controls a variable circular aperture that, by opening and closing, adjusts the amount of light falling on the retina. This aperture, known as the pupil, is seen as a black circle in the centre of the iris. In dim light the pupil opens wide to let through as much light as possible while in very bright conditions it closes up to reduce the amount of light entering the eye.
1.2. The Photoreceptors
Having formed an image on the retina, the information describing it must be transformed into electrical signals that are transmitted to the brain. The first stage of this process is done by photoreceptors distributed all over the retina. There are about 150 million of them, distributed most densely in the centre of the field of view in the region shown as the fovea in Fig. 1.2, with the density falling off with distance from the fovea. These photoreceptors are of two kinds. One kind consists of long thin receptors called rods, which are very sensitive and can record very low light levels. They give what is known as scotopic vision, which would, for example, operate at night with the Moon as illumination. The second kind of receptor is stubbier; they are called cones and they are responsible for photopic vision, which occurs under everyday normal light conditions.
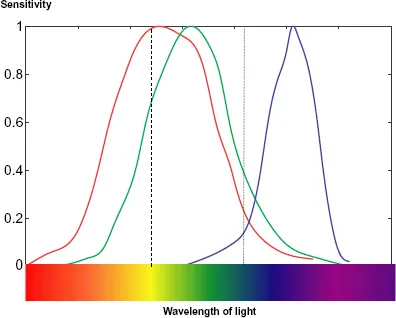
Apart from their sensitivity, rods and cones have another important difference — in the degree to which they give the perception of colour. There is no perception of colour with scotopic vision — just shades of grey, although it sometimes seems that moonlight has a bluish tinge. By contrast, photopic vision is fully chromatic and this is because there are three kinds of cones with different colour responses, illustrated in Fig. 1.3.
In every part of the retina there is a mixture of the different types of cone and the relative responses of the three types gives the colour perceived. For example, if the red response is somewhat greater than the green response, with virtually no blue response, as indicated by the dashed line in Fig. 1.3, then the colour seen will be yellow. As another example, if the largest contribution is from green with a smaller contribution from red and a significant contribution from blue, as indicated by the dotted line in the figure, then the colour seen will be cyan (green-blue). In practice the light received by the eye normally covers a range of wavelengths but the relative response of the three kinds of receptor will still indicate the colour perceived. If all the receptors are affected equally then the colour seen is white. Usually the relative responses of the three kinds of receptor correspond to that from some wavelength of the spectrum plus equal responses corresponding to white light, in which case the colour seen is an unsaturated spectral colour — sometimes known as a pastel shade. Finally, if the responses are strong for blue and red but weaker for green then this corresponds to no colour in the spectrum and gives the colour sensation of purple.
Figure 1.3 The relative sensitivity for each type of cone in the retina, each normalized to a maximum of unity.
The strength of the responses of the three kinds of receptor in terms of their output do not have the same maximum value, as indicated in Fig. 1.3, but the essentials of the process of colour perception are as described.
The foveal region contains only cones, which are tightly packed there, hence giving the greatest acuity of vision at the centre of the field of view. Moving outwards from the fovea, the proportion of rods becomes greater, giving better peripheral vision under low light conditions but with deterioration of acuity as the density of photoreceptors is lower. The responses of individual rods and cones influence the electrical signals that travel along individual optic nerve fibres that pass into the visual cortex in the brain. However, there is not a oneto-one relationship between photoreceptors and optic nerve fibres; the responses of about 150 million receptors are funnelled into about 1 million optic nerve fibres. This funnelling process is carried out by four intermediate layers of specialist cells that mix and process the data from the photoreceptors before they reach the optic nerve fibres. There is no complete understanding of the way this system operates but in order to appreciate the processes that occur we first need to know something about the way that nerve cells work and communicate with each other.
1.3. The Way that Nerve Cells Operate and Communicate
The behaviour of all nervous systems, including that of the eye, is extremely complex and involves physical processes triggered by various chemical agents. Here we give just a very elementary description of the action of a typical nerve cell, sufficient for the purpose of describing the general processes that occur in the neural networks of the eye.
Figure 1.4 shows a schematic nerve cell, or neuron. At one end there are dendrites, connections that receive information from other neurons. At the other end is the axon, a channel along which the electrical nerve impulse travels and the terminus of which communicates with other neurons. The interior of the cell contains an excess of potassium ions, K+, potassium atoms lacking one electron, which gives it a positive charge. Negatively charged molecules within the cell counterbalance these positive charges to some extent. Outside the cell there is an excess of sodium ions, Na+, and in the resting state the electric potential outside the cell is greater than that inside the cell. The ions are able to permeate through the membrane that acts as a sheath for the cell but there are chemicals within the membrane that act like ion pumps, a separate one for each ion, so maintaining the relative concentrations of the two kinds of ion. A cell in this situation is said to be polarized.
Figure 1.4 A schematic nerve cell.
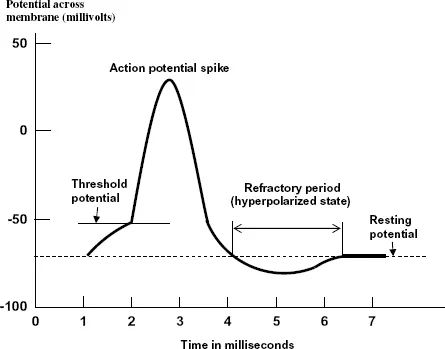
The connections between axons and dendrites are called synapses and for all cells, except those in the brain, the transmission of the signal across a synapse is by chemicals known as neurotransmitters. These traverse the tiny synaptic gaps and enter the receptor cell via its dendrites where they become attached to chemical receptors. Synapses can either excite activity or inhibit activity in the receptor cell dependent on what happens when the neurotransmitter arrives. Activity is stimulated when the neurotransmitter opens up sodium channels so that more sodium ions enter the cell. When this happens the potential difference between the inside and outside will reverse — the cell is then depolarized and an electrical impulse is transmitted through the neuron. This occurs when the depolarization potential difference reaches a certain critical level — the threshold potential. As sodium channels are opened up in one part of the cell so the potential difference reaches a critical level to open up sodium channels further along the cell and this way, in a few milliseconds, the electrical pulse traverses the neuron, moving towards the end of the axon. When the electric pulse arrives at the terminus of the axon, the membrane there allows the passage of doubly charged calcium ions, Ca2+, into the cell and this triggers the release of the neurotransmitter towards the dendrites of the next cell. The form of the action potential, i.e. the potential difference across the membrane, at each point of the neuron as a function of time is shown in Fig. 1.5. As more and more sodium ions enter the cell so this stimulates the potassium channels to open and potassium ions move to the outside of the cell until eventually the cell returns towards its polarized state with a slight overreaction to become hyperpolarized, where the difference between the outside and inside potential is higher than the normal resting state of the neuron. When the cell is hyperpolarized the sodium and potassium pumps return to their normal mode of action and restore the condition in which there is an excess of sodium ions on the outside and an excess of potassium ions on the inside. While this state is being restored the neuron cannot be stimulated; it is then in its refractory period.
The rate at which action potential spikes are generated depends on the strength of the stimulation but at a certain level of stimulation a stage of saturation is reached and no greater rate of spike discharge is then possible, no matter how strong the stimulation. The neuron reaction we have just described is when it is excited by the oncoming neurotransmitter. However, there can also be an inhibitory response of the cell, when it is the potassium rather than the sodium channels that are stimulated to open. In this case potassium ions flood out of the cell, it becomes hyperpolarized, and the impulse is stopped dead in its tracks through the system. Both excitatory and inhibitory reactions are important in the functioning of the optical neural network system.
Figure 1.5 The action potential of a neuron. The potential across the membrane is the potential inside minus the potential outside and hence is negative in the resting state.
1.4. The Neural Network of the Eye
There are four layers of neural cells betw...