![]()
1
Introduction
The honey bee exhibits a combination of individual traits and social cooperation which is unparalleled in the animal kingdom. The multiple levels at which the honey bee expresses adaptations to its world provide one of the richest sources for study and knowledge among all organisms, made even more enriching by the economic benefits the honey bee provides. The honey bee can be, and has been, studied from perspectives as divergent as those of a beekeeper and a molecular biologist, a behavioral ecologist and a primitive honey hunter, a student of social behavior and a doctor interested in allergic reactions, and all have contributed immensely to our understanding of this most well-studied insect.
A quick glimpse inside the nest makes it readily apparent why honey bees have fascinated us from the earliest days of scientific observation (Fig. 1.1). The infrastructure of the nest, the exquisitely uniform and functional comb, is composed of beeswax produced by the workers and constructed into a repeating series of almost perfect hexagonal cells. The comb provides the substrate for interactions between colony members and is used for almost everything imaginable, from larval nursery to pantry to message center. As the stage for colony activities, it provides us with a rich arena for observing the individual and social behaviors that are at the heart of honey bee society.
At the individual level, honey bees have not one but three types of colony members: queens, drones, and workers, each with their own specializations and place in honey bee society. The aptly named queen reigns over the nest, surrounded by attendants and fed the rich food she requires to perform her few but crucial tasks in the colony. Her slim lines hide the huge ovaries which make her an extraordinary egg-laying machine, capable of laying thousands of eggs a day, and her calm behavior masks her powerful pheromones, chemical signals to recipient workers which control many of their behaviors and provide part of the social glue which holds honey bee life together.
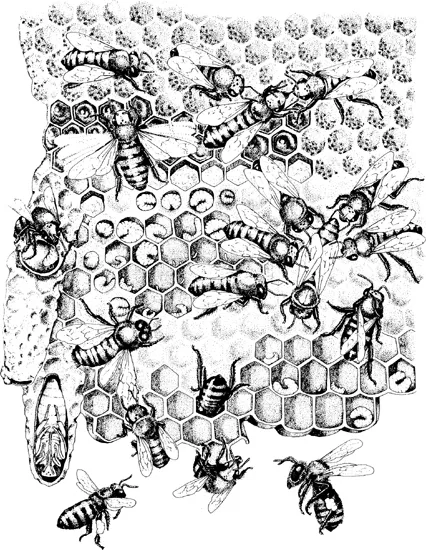
Fig. 1.1
A typical scene from a honey bee colony during the summer. Cells in the top section of the comb contain capped honey and uncapped nectar and pollen, while the middle and bottom cells contain eggs, larvae, and capped brood. The elongated cells on the left side of the comb are queen cells with various stages of developing queen brood inside; the bottom queen cell is cutaway to show a queen almost ready to emerge. The adult workers are, in clockwise order from the top left: resting, fanning to evaporate water from nectar, exchanging food, attending the queen in a retinue surrounding her, grooming, returning to the nest with nectar and pollen, building the comb, inspecting cells, driving a drone out of the nest, and capping a queen cell. (Based on Wilson, 1971.)
The other sexual members of this society, the drones, are tended and fed by the workers, although they perform only one function, the all-important one of mating the queen, after which they die. With their large eyes, flight muscles, and powerful mating urge, the drones are magnificently constructed for this one task.
The workers perform endless and diverse tasks in the nest, sometimes dying from stinging their colony’s opponents, and rarely reproducing. At any one time workers may be found walking on the comb surface, perhaps tending the brood, cleaning debris from the nest, capping cells, ripening or storing honey, organizing pollen for storage near the brood, feeding or grooming the queen, or any one of a myriad other activities.
Not only have the complex individual behaviors manifested by these three castes prompted our investigations into honey bee life, but their sociality and adaptability has fueled the study of this insect to an extent not found for many other organisms on earth. The social nature of this organism, its tremendous ability to regulate its functions as a colony of individuals according to events within and outside the nest, provides the key to its success and makes the contribution of individuals much greater than the sum of their individual behaviors. We are further motivated to study this insect by an economic stake, since the products of the hive are crucially important to our agricultural systems. Not only do honey bees provide us with honey, wax, propolis, royal jelly, and pollen, but they also pollinate a good portion of our crops, including such diverse agricultural plants as fruit trees, oilseeds, small berries, and forage crops.
The study of honey bees is not new; extant cave paintings dating back many thousands of years show honey bee nests in marvelous detail. Certainly these are the forerunners of the thousands of articles written in the scientific and popular journals of contemporary science. It is an intangible quality of honey bee society that draws us to its study, an almost mystical presence which pervades their nests and can only be partly dissected, analyzed, and reported on by its observers. This book is an attempt to provide at least a glimpse into the world of this queen of insects.
![]()
2
The Origin and Evolutionary History of Bees
Bees are essentially wasps which abandoned predation in favor of provisioning nests with nectar and pollen. Most of the aculeate (stinging) wasps which are related to bees prey on other insects or spiders for larval food. However, adults frequently feed on nectar from flowers, and their mouthparts are often well adapted for sucking or lapping. Bees are thought to have evolved from a wasp ancestor, probably a sphecid, with mouthparts capable of ingesting nectar, which began collecting pollen to feed to their brood instead of killing prey. While bees have diverged from wasps in many characteristics (Michener, 1974), the most distinctive morphological differences involve specializations for pollen collection. All bees have at least a few plumose hairs and broadened hind legs, both adaptations for gathering pollen and transporting it back to the nest. Because of their distinctive pollen-collecting structures and habits, bees are classified in their own superfamily, Apoidea—order: Hymenoptera—(Culliney, 1983), although Michener (1974) has proposed returning to an older system which included the sphecoid wasps in the same superfamily as the bees.
The earliest bees may have arisen in the xeric interior of the paleocontinent Gondwana, which was probably the area of origin for flowering angiosperm plants (Raven and Axelrod, 1974). Although the fossil record of bees is far from complete, they are thought to have diverged from the sphecoid wasps during the middle Cretaceous period, about 100 million years ago (Michener, 1974), coincidental with the appearance of angiosperms as the dominant vegetation. The earliest known fossil bees are from the Eocene period 40 million years ago (Manning, 1952; Kelner-Pillault, 1969; Zeuner and Manning, 1976), but since these specimens were already highly specialized it is clear that bees arose much earlier. At any rate, the evolution and divergence of bees has been closely linked to that of the angiosperm plants, with the plants evolving flowers with odors, shapes, colors, and excess nectar and pollen food rewards to attract the bees, and the bees in turn providing a mechanism to transfer pollen between plants. The coevolution of these two groups has been one of the dominant themes of recent evolutionary history.
There are currently 10 or 11 families of bees (Michener, 1974; Michener and Greenberg, 1980), with approximately 700 genera (Malyshev, 1968) and 20,000 living species (Michener, 1969). These can be divided into two major groups, the more primitive short-tongued bees and the more advanced long-tongued bees (Fig. 2.1). The short-tongued bees probably utilized the shallow flowers characteristic of the earliest angiosperms, but some bees evolved longer mouthparts as many of the angiosperms developed longer, tubular flowers. These adaptations allowed the long-tongued bees to take advantage of the increasing complexity of advanced angiosperm flowers.
The honey bee Apis mellifera L. was one of these long-tongued bee species. Its scientific name means “honey-bearing or honey-producing bee,” and refers to the bees’ habit of collecting nectar and producing from it copious amounts of honey to allow colonies to survive dearth periods.
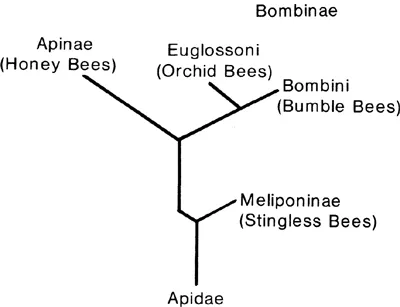
Honey bees are classified in the family Apidae, and their close relatives include the orchid bees (Euglossini), the bumble bees (Bombini), and the stingless bees (Meliponinae) (Winston and Michener, 1977; Kimsey, 1984) (Fig. 2.2). All of the Apidae are characterized by the presence of a corbicula or pollen basket on the outer surface of each hind tibia, at least in workers, and this structure is used to carry pollen and nest-building materials. Pollen is generally pushed up into the corbicula from the distal end of the tibia, and this loading mechanism is also a distinctive feature of the apids (Winston and Michener, 1977; Michener, Winston, and Jander, 1978). All of the Apidae show some degree of social behavior, and the Meliponinae and Apinae have the most elaborate social behavior of all the bees. Reviews of the natural history and biology of stingless bees and orchid bees can be found in Michener (1974), while Free and Butler (1959), Alford (1975), and Heinrich (1979a) have written accounts of bumble bee societies.
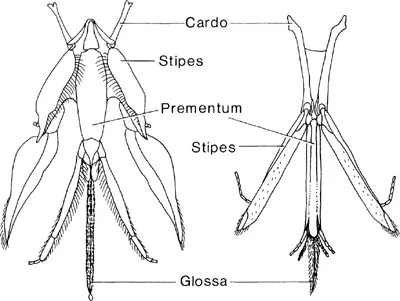
Fig. 2.1
Mouthparts of representative long-tongued (left) and short-tongued (right) bees. (Redrawn from Michener, 1974.)
Fig. 2.2
Taxonomic relationships between bees in the family Apidae.
Honey Bee Origins
The modern honey bees (Apidae: Apini) are all classified in only one genus, Apis, which includes five species: the common honey bee A. mellifera, the giant honey bees A. dorsata and A. laboriosa, the Indian honey bee A. cerana, and the dwarf honey bee A. florea. The earliest Apini fossil specimens have been found in Baltic Amber from Eocene layers approximately 40 million years old, and these ancient but extinct apines have been classified in their own genus Electrapis (Manning, 1960; Zeuner and Manning, 1976; Culliney, 1983). A fossilized honey bee comb has recently been discovered in Malaysia dating from the late Tertiary or early Quaternary period, suggesting an earlier origin of the genus (Stauffer, 1979). This finding and the fact that fossil honey bee specimens are generally found with individuals grouped together suggest early evolution of social behavior in the Apini. Rapid evolution during the next 10 million years is shown by specimens found from the Oligocene, when considerable change occurred in external morphology. Also, comparative biochemical studies of extant bees have indicated a greater degree of amino acid substitution in A. mellifera compared to other bees, and therefore a more rapid protein evolutionary rate in the honey bee lineage than for other bees (Carlson and Brosemer, 1971, 1973). On the basis of morphological evidence, however, there has been relatively little change in honey bees during the last 30 million years (Culliney, 1983), and the physical resemblance of fossil forms to modern worker bees suggests that complex social behavior had already developed by the Miocene, 27 million years ago.
The natural geographic distribution of the genus Apis shows its greatest species diversity in India and adjacent regions, with all of the species except A. mellifera found there. Therefore, these regions probably constitute the area of origin and early evolution of the Apini (Doediker, Thakar, and Shaw, 1959; Michener, 1974; Doediker, 1978). A. mellifera is thought to have originated in the African tropics or subtropics during the Tertiary period, migrating to western Asia and colder European climates somewhat later. Until modern times Apis was not found anywhere in the western hemisphere, Australia, or the Pacific except for some of the continental islands such as Japan, Formosa, the Philippines, and Indonesia (Michener, 1974). But movement of bees by European settlers for beekeeping has resulted in A. mellifera now having worldwide distribution and some of the other species being more widespread in Asia.
The dwarf honey bee A. florea appears to maintain several ancestral characteristics of the genus Apis and is probably the closest living descendant of the earliest honey bees. Workers are small, approximately 7 mm in length, and colonies construct a single comb supported from branches, frequently at sites surrounded by dense vegetation (Seeley, Seeley and Akratanakul, 1982). Their communicative dances occur on a horizontal platform built on the top of the comb, and thus direction to flowers is indicated by straight runs toward the food source. Colonies tend to be small, less than 5000 individuals, and the workers are relatively docile (Michener, 1974).
Two other closely related species of honey bees also construct nests consisting of a single comb in the open, the giant honey bees A. dorsata and A. laboriosa. These are large, feisty bees 17–19 mm in length, with 20,000 or more workers in a colony. Their nests are constructed high in trees or suspended from open cliff faces, and nests do not need to be concealed because of the worker’s aggressive nature. Nests are also frequently aggregated, and colonies may migrate up and down mountains to take advantage of seasonal nectar sources. Communicative dances are more advanced than for A. florea since they occur on the vertical comb face, and the direction to flowers has to be translated by the workers from the vertical dance angle to the direction of the sun (Michener, 1974). A. laboriosa is the larger of the two species, and its large size, dark color, and long hair coat are probably adaptations for its high-altitude Himalayan habitat (Sakagami, Matsumura, and Ito, 1980).
The other two honey bee species, A. cerana and A. mellifera, are medium sized bees (10–11 mm) which generally build multiple comb nests inside cavities. Colonies of A. cerana are relatively small, 6000–7000 workers (Seeley, Seeley, and Akratanakul, 1...