![]()
Chapter 1. Botanical and Marine Oils for Treatment of Arthritis
Abundant experimental evidence supports the view that prostaglandins, thromboxanes, and leukotrienes, collectively termed eicosanoids, participate in development and regulation of immunological and inflammatory responses. Because essential fatty acids are precursors to eicosanoids, and because essential fatty acids are important determinants of cell membrane structure and function, they influence immune responses. Diseases such as rheumatoid arthritis (RA), and systemic lupus erythematosus (SLE), characterized by abnormal immune responses, persistent inflammation, and tissue injury, may therefore be amenable to control by dietary means. Indeed, considerable public interest has been provoked recently by treatment of a variety of disorders, including RA, with plant seed and fish oils. The established and potential uses of unsaturated fatty acids for treatment of inflammatory arthritis and related conditions are examined in this chapter.
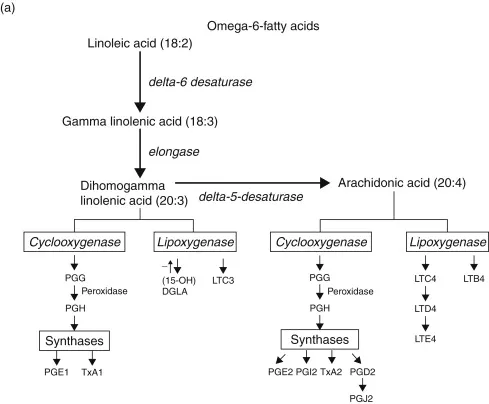
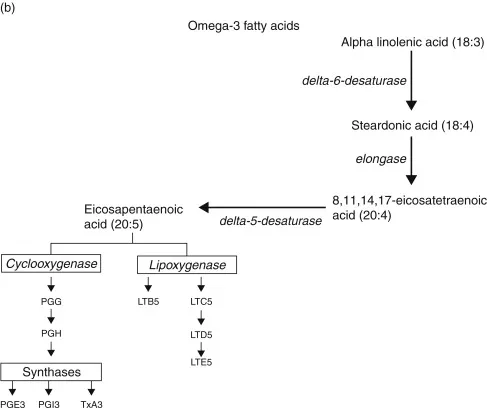
Essential fatty acids are “essential” [1] not only because of their physiologic importance, but because they must be derived in either full or partially elaborated form from the diet. Thus, these acids may be classified as vitamins (indeed, they were once called vitamin F). Two groups of fatty acids are essential to the body: the omega-6 (n6) series, derived from linoleic acid (18:2 n6) and the omega-3 series, derived from α-linolenic acid (18:3 n3). In these notations, 18 is the number of carbon atoms in a molecule, the second number is the number of double carbon–carbon bonds (degree of unsaturation), and the number after the “n” is the position of the first double bond starting from the methyl (omega) end of the fatty acid chain. The metabolic sequences of the two fatty acid series are shown in Fig. 1.1. Fatty acids provide energy, are an integral part of cell membranes, and are precursors for the eicosanoids. Alteration of the eicosanoid profile by administration of fatty acid precursors other than arachidonic acid is one approach to modulation of host defense.
FIGURE 1.1. Metabolic pathways for omega-6 and omega-3 fatty acids. During inflammatory responses, cells are activated to release arachidonic acid or EPA. Activity of the bifunctional enzyme COX oxidizes the precursor fatty acid to PGG and a subsequent peroxidase reaction yields PGH. Then under the influence of terminal synthases, prostaglandins, thromboxanes, and prostacyclins are formed.
Although changes in eicosanoid production due to alteration of fatty acid intake formed the basis of the current hypothesis for antiinflammatory and immunoregulatory effects of this type of treatment, it is likely that the precursor fatty acids themselves influence immune responses. Studies in animals and humans have shown that changes in essential fatty acid intake alter the fatty acid composition of cell membranes [2]. For example, in essential fatty acid deficiency, deprivation of linoleic acid leads to deficiency of arachidonic acid and impairment of prostaglandin synthesis. Essential fatty acid deficiency causes many pathologic changes, but it also reduces the severity of inflammation in experimental animal models. Fasting also has a salutary effect on symptoms of patients with RA [3]. Because neither induction of essential fatty acid deficiency nor fasting are likely to be popular treatments, it follows that it would be more prudent to modify or supplement, rather than delete, lipid intake.
The extraordinary rapidity with which platelets adhere to damaged tissue, aggregate, and release potent biologically active materials, suggests that the platelet is well suited to be a cellular trigger for the inflammatory response [4]. Thus, experiments directed at suppression of thromboxane synthesis, enhancement of prostacyclin production, and inhibition of platelet aggregation, have been conducted in an effort to limit inflammatory responses. Fish oil lipids, rich in eicosapentaenoic acid (EPA; 20:5 n3), inhibit formation of the cyclooxygenase (COX) products thromboxane A2 (TXA2) and prostaglandin E2 (PGE2) derived from arachidonic acid. The newly formed thromboxane A3, has much less ability than TXA2 to constrict blood vessels and aggregate platelets. In addition, production of prostaglandin I2 (PGI2, prostacyclin) by endothelial cells is not reduced appreciably by increased EPA content, and the physiologic activity of newly synthesized PGI3 is added to that of PGI2[5]. Moreover, diets enriched in fish oil reduce production of the potent mediator of inflammation leukotriene B4 (LTB4) via 5-lipoxygenase in stimulated neutrophils and monocytes, and induce the appearance of EPA-derived LTB5, which is far less biologically active than LTB4[6, 7].
Evidence obtained from experiments in vitro and in vivo in small animals and in humans suggests that other novel fatty acids may be safe and effective antiinflammatory and immunomodulatory agents. For example, certain botanical lipids, notably those extracted from seeds of the evening primrose and borage plants, contain relatively large amounts of gammalinolenic acid (GLA; 18:3 n6). This fatty acid is converted rapidly to dihomogammalinolenic acid (DGLA; 20:3 n6), the fatty acid precursor of the monoenoic prostaglandins, such as prostaglandin E1 (PGE1). In humans, the delta-5-desaturase that converts DGLA to arachidonic acid is sluggish. Thus, concentrations of arachidonate do not increase appreciably in patients treated long term with GLA. DGLA competes with arachidonate for oxidative enzymes, thereby reducing synthesis of COX products derived from arachidonate. In addition, DGLA cannot be converted to inflammatory leukotrienes by 5-lipoxygenase. Instead, it is converted to 15-hydroxy-DGLA (Fig. 1.1), which has the additional virtue of suppressing 5-lipoxygenase activity [8].
Enrichment with DGLA of human synovial cells (which line the joint space, and proliferation of which leads to cartilage degradation and bone erosion in patients with RA) in culture leads to a marked reduction of PGE2 synthesis, a substantial increase in PGE1 production, and a reduction in interleukin-1β (IL-1β) induced synovial cell proliferation. Addition to cultures of arachidonic acid (which increases PGE2 substantially) or EPA does not modify synovial cell proliferation. The antiproliferative effect of DGLA is prevented by indomethacin [9]. Although PGE compounds are clearly local mediators of inflammation [10], evidence from both in vitro and in vivo experiments indicates that they can also suppress diverse effector systems of inflammation [11, 12]. In addition, PGE can both enhance and diminish cellular and humoral immune responses, observations that reinforce a view of these compounds as regulators of cell function. Defective regulation of inflammatory responses, or disordered immune mechanisms, or both, are probably central to the pathologic processes encountered in rheumatic diseases such as RA and SLE. PGE1, largely by virtue of its ability to influence cellular cyclic nucleo-tides and signal transduction pathways, appears to be an important regulator of cell function and, therefore, potentially able to influence the function of those cells that participate in tissue injury in these diseases. A regulatory effect of PGE is not without precedent. For example, PGE inhibits the release of noradrenaline from the spleen in response to sympathetic nerve stimulation, and PGE is released from the spleen when it contracts in response to sympathetic nerve stimulation [13]. Thus, by a feedback mechanism, the contracting smooth muscle can reduce the stimulus that is leading to its contraction. PGE release may therefore be a defense mechanism aimed at minimizing potential injury. Certainly PGE compounds serve to protect gastric mucosa and kidney function. Suppression by PGE1 of leukocyte effector functions and of mediator release suggests that PGE1 has antiinflammatory effects.
The role of prostaglandins in the inflammatory process is not as well defined as once supposed, because the stable prostaglandins PGE and PGI2 have antiinflammatory and inflammatory effects [14]. PGJ and lipoxins appear to protect against runaway inflammatory responses. Even LTB4 appears capable of modulating inflammation and immune responses [15]. The observation that PGE1 inhibits platelet aggregation led to the notion that COX products of arachidonic acid metabolism might have antiinflammatory activity. As it becomes more clear that nonsteroidal antiinflammatory drugs (NSAIDS) have antiinflammatory effects other than interference with COX production and subsequent prostaglandin inhibition [16], the potential protective effects of prostaglandins are being considered.
Other eicosanoids also serve to counter inflammation: PGJ2, formed from the dehydration of PGD2, appears to function as a brake on the inflammatory response [17]. Lipoxins are a new class of eicosanoids, which arise from the sequential action of 5- and 15-lipoxygenases. They serve as stop signals in that they prevent leukocyte-mediated tissue injury, thereby facilitating resolution of inflammation [18]. Human endothelial cells treated with aspirin in vitro convert EPA to antiinflammatory lipoxins. These novel compounds are also found in inflammatory exudates from animals administered aspirin and fish oil [19].
Thus, it will take exquisitely selective alteration of particular points in the arachidonic acid cascade (as prostaglandin and thromboxane synthases, or eicosanoid receptors) to modify it to the advantage of the host. The selective COX-2 inhibitors were an attempt to do that. It is clear that these agents are equivalent in efficacy to the traditional NSAIDs and that they reduce the more serious gastrointestinal events (ulcer formation, bleeding, and perforation). However, COX-2 is upregulated in stimulated endothelial cells [20]. Thus, selective COX-2 inhibition results in suppression of PGI2 synthesis by these cells. Because platelets lack COX-2, platelet thromboxane production is spared by selective COX-2 inhibition, and the platelet–blood vessel wall balance mediated between the effects of thromboxane (platelet aggregation, vasoconstriction) and PGI2 (inhibition of platelet aggregation, vasodilation) is tilted toward promotion of thrombosis. These biochemical truths did indeed translate to the clinical findings that selective COX-2 inhibition is associated with an increased incidence of acute myocardial infarction (coronary artery thrombosis) [21].Thus, botanical and marine oils, which protect the gastric mucosa and counter thrombosis, should find a place in the armamentarium of anti-inflammatory agents.
PGE1 has remained a bit of an orphan among the eicosanoids, mainly because of a long-held notion that not enough is produced by human cells to be of use, and that its biological effects are no different from those of PGE2 and PGI2. Contrary to popular belief, however, PGE1 is found in physiologically important amounts in humans. Lost in the vast literature on the “arachidonic acid cascade” are the early observations of Bygdeman and Samuelsson [22], who found (using bioassay) the concentration of PGE1 in human seminal plasma (16 μg/ml) to be higher than PGE2 (13 μg/ml), PGE3 (3 μg/ml), PGF1α (2 μg/ml), and PGF2α (12 μg/ml). Furthermore, Karim et al. [23] found PGE1 to be the sole PGE in human thymus. Prostaglandin immunoassays usually do not distinguish between PGE1 and PGE2. To identify PGE1, PGs must first be separated by thin layer or high performance l...