Introduces all of the essential cell biology and developmental biology background for the study of stem cells
This book gives you all the important information you need to become a stem cell scientist. It covers the characterization of cells, genetic techniques for modifying cells and organisms, tissue culture technology, transplantation immunology, properties of pluripotent and tissue specific stem cells and, in particular, the relevant aspects of mammalian developmental biology. It dispels many misconceptions about stem cells—especially that they can be miracle cells that can cure all ills. The book puts emphasis on stem cell behavior in its biological context and on how to study it. Throughout, the approach is simple, direct, and logical, and evidence is given to support conclusions.
Stem cell biology has huge potential for advancing therapies for many distressing and recalcitrant diseases, and its potential will be realized most quickly when as many people as possible have a good grounding in the science of stem cells.
Content focused on the basic science underpinning stem cell biology
Covers techniques of studying cell properties and cell lineage in vivo and in vitro
Explains the basics of embryonic development and cell differentiation, as well as the essential cell biology processes of signaling, gene expression, and cell division
Includes instructor resources such as further reading and figures for downloading
Offers an online supplement summarizing current clinical applications of stem cells
Written by a prominent leader in the field, The Science of Stem Cells is an ideal course book for advanced undergraduates or graduate students studying stem cell biology, regenerative medicine, tissue engineering, and other topics of science and biology.
Preguntas frecuentes
¿Cómo cancelo mi suscripción?
Simplemente, dirígete a la sección ajustes de la cuenta y haz clic en «Cancelar suscripción». Así de sencillo. Después de cancelar tu suscripción, esta permanecerá activa el tiempo restante que hayas pagado. Obtén más información aquí.
¿Cómo descargo los libros?
Por el momento, todos nuestros libros ePub adaptables a dispositivos móviles se pueden descargar a través de la aplicación. La mayor parte de nuestros PDF también se puede descargar y ya estamos trabajando para que el resto también sea descargable. Obtén más información aquí.
¿En qué se diferencian los planes de precios?
Ambos planes te permiten acceder por completo a la biblioteca y a todas las funciones de Perlego. Las únicas diferencias son el precio y el período de suscripción: con el plan anual ahorrarás en torno a un 30 % en comparación con 12 meses de un plan mensual.
¿Qué es Perlego?
Somos un servicio de suscripción de libros de texto en línea que te permite acceder a toda una biblioteca en línea por menos de lo que cuesta un libro al mes. Con más de un millón de libros sobre más de 1000 categorías, ¡tenemos todo lo que necesitas! Obtén más información aquí.
¿Perlego ofrece la función de texto a voz?
Busca el símbolo de lectura en voz alta en tu próximo libro para ver si puedes escucharlo. La herramienta de lectura en voz alta lee el texto en voz alta por ti, resaltando el texto a medida que se lee. Puedes pausarla, acelerarla y ralentizarla. Obtén más información aquí.
¿Es The Science of Stem Cells un PDF/ePUB en línea?
Sí, puedes acceder a The Science of Stem Cells de Jonathan M. W. Slack en formato PDF o ePUB, así como a otros libros populares de Ciencias biológicas y Biología celular. Tenemos más de un millón de libros disponibles en nuestro catálogo para que explores.
In the popular media and even in some medical circles, stem cells are presented as miracle cells that can do anything. When administered to a patient with some serious disease they will rebuild the damaged tissues and make the patient young again. Alas, in reality there are no such cells. However, there are cells that exhibit stem cell behavior and the future of regenerative medicine will undoubtedly be built on a good scientific understanding of their properties. In this chapter these properties are briefly outlined, and in the remainder of the book each of them will be underpinned by an explanation of the relevant areas of science and technology.
A list of characteristics of stem cell behavior that is generally agreed upon is the following:
Stem cells reproduce themselves.
Stem cells generate progeny destined to differentiate into functional cell types.
Stem cells persist for a long time.
Stem cell behavior is regulated by the immediate environment (the niche).
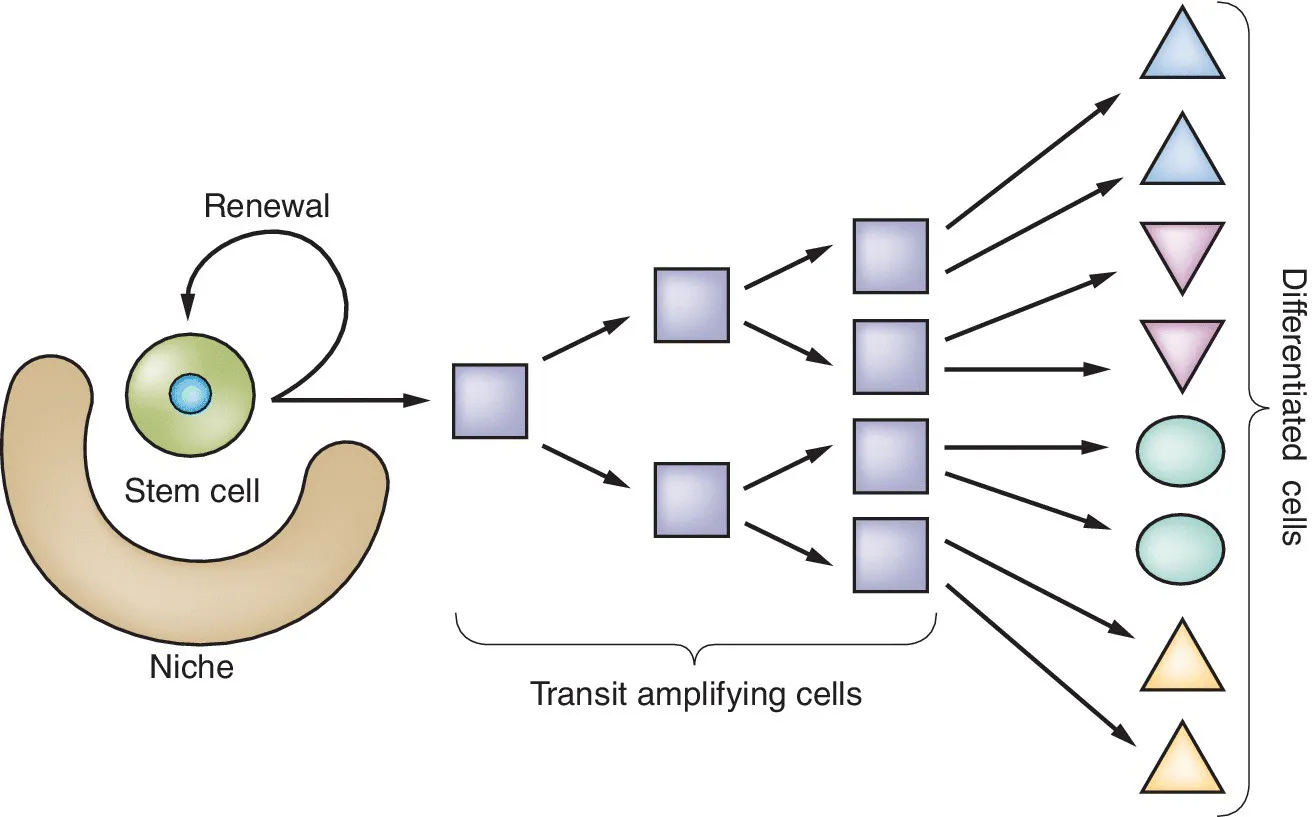
This is shown diagrammatically in Figure 1.1. The first two items on the list indicate the key abilities of self‐renewal and of generation of differentiated progeny. As will be explained below, these abilities may be shown at a cell population level rather than by every single stem cell at every one of its divisions. Also, the second item indicates “destined to differentiate” meaning that cell division may continue for a while before differentiation, but not indefinitely. Cells derived from stem cells that proliferate for a limited number of cycles are called progenitor cells or transit amplifying cells. The third item on the list means that if the stem cell population is one of those that exists in tissue culture then it should be capable of indefinite growth, while if it is part of an organism it should be very long lasting, normally persisting for the whole life of the organism. The fourth characteristic indicates that all stem cells exist in a specific micro‐environment that controls their program of division and differentiation. This may seem at first sight only to apply to stem cells within the body and not to those grown in vitro, but in order to get them to grow, the cells in vitro are always provided with specialized medium ingredients that, in effect, mimic the components normally provided in the niche.
Figure 1.1 A consensus diagram showing stem cell behavior.
(Modified from Slack, J.M.W. (2013). Essential Developmental Biology, 3rd edn. Reproduced with the permission of John Wiley and Sons.)
This fourfold definition involves not just intrinsic properties of stem cells, but also properties that depend on aspects of their environment such as the lifespan of the animal, the nature of the niche, or the composition of the culture medium. This emphasizes the fact that the goal of stem cell biology is understanding the behavior and not just the intrinsic nature of stem cells. To achieve this, the characteristics of the stem cell environment are just as important as the properties of the stem cells themselves. Moreover, understanding stem cell behavior means understanding various aspects of cell and developmental biology which are not always familiar to workers in stem cell laboratories.
The above definition is of value in indicating the special characteristics of stem cell behavior, but is also helpful in indicating what is not stem cell behavior. For example, most of the cells in the body that are dividing are not stem cells. In particular cells in the embryo that differentiate after a certain period of time, such as the earliest cells formed by division of the fertilized egg, are not stem cells. Nor are differentiated cells that divide during postnatal life to generate more of themselves, such as hepatocytes or tissue‐resident macrophages. A common term found in the literature is “stem/progenitor cell”. This is a singularly unhelpful designation as it conflates two entirely different cell behaviors. Progenitor cells are precisely those that differentiate into functional cell types after a finite period of multiplication. They include the transit amplifying cells that arise from stem cells (Figure 1.1) and also cells of the embryo and of the growing individual that are destined to differentiate after a certain time.
Real stem cells comprise two fundamentally different types: pluripotent stem cells that exist only in vitro, and tissue‐specific stem cells that exist in vivo in the postnatal organism. Pluripotent stem cells comprise embryonic stem cells (ESC) and induced pluripotent stem cells (iPSC). There are various subdivisions that will be considered later, but the essential features of these cells are first that they can be propagated without limit in vitro, and second that, under appropriate culture conditions, they are able to give rise to a wide variety of cell types, perhaps all the cell types in the normal organism except for the trophectoderm of the placenta. By contrast, tissue‐specific stem cells exist within the body and generate progeny to repopulate the tissue in question. Well‐studied tissue‐specific stem cells include those of the hematopoietic (blood‐forming) system, the epidermis, the intestinal epithelium and the spermatogonia of the testis. Under normal circumstances, tissue‐specific stem cells do not produce cells characteristic of other tissue types. There are also some well‐characterized stem cells that do not undergo continuous division, but seem to be kept in reserve to deal with tissue regeneration when required. A good example is the muscle satellite cells, which are normally quiescent but are able to be mobilized to divide and fuse to form new myofibers following injury. This type of stem cell behavior is sometimes called facultative.
Many criteria for identifying stem cells have been proposed and used. These are briefly listed here and the concepts and technologies will be developed in later chapters of the book.
Stem Cell Markers
Very often a cell is said to be a stem cell because it expresses one or more gene products associated with stem cells. However, there is no molecular marker that identifies all stem cells and excludes all non‐stem cells. Those components required for general cell metabolism and cell division are certainly found in all stem cells, but they are also found in many other cell populations as well.
Pluripotent stem cells (ESC and iPSC) express an important network of transcription factors which are necessary for maintenance of the pluripotent state (see Chapter 6). Transcription factors are the class of proteins that control the expression of specific genes. A key member of the pluripotency group is the POU‐domain transcription factor OCT4 (also known as OCT3 and POU5F1). The presence of OCT4 is certainly necessary for the properties of pluripotent stem cells. However it is not expressed in any type of tissue‐specific stem cells except at a low level in spermatogonia.
A component that might be expected to be found in all stem cells is the telomerase complex. At the end of each chromosome is a structure called the telomere, made up in vertebrate animals of many repeats of the simple sequence TTAGGG. Because of the nature of DNA replication, the double helix cannot be copied right up to the end, so a part of the telomere is lost in each cell cycle. After enough cycles, the erosion of chromosome ends activates the system which senses DNA double‐stranded breaks and causes death of the cell. This process is an important reason for the limited survival time of most primary tissue culture cell lines, which undergo senescence after a certain number of population doublings in vitro. Obviously there must be a mechanism for repairing telomeres in vivo, and this is provided by the telomerase complex, of which the most important components are an RNA‐dependent DNA polymerase called TERT, and an RNA called TERC which contains the template CCCTAA for the telomere sequence. High levels of telomerase are found in germ cells, ensuring the survival of full length chromosomes for the next generation. Telomerase is also upregulated in permanent (“transformed”) tissue culture cell lines and in most cancers. However most types of somatic cell have little or no telomerase. Tissue‐specific stem cells do contain some telomerase; generally enough to maintain cell division for a normal lifetime, but not enough to fully reverse the erosion of the telomeres. In situations such as repeated transplantation of hematopoietic stem cells from one mouse to another, there is an upper limit to the number of possible transplants and this is determined at least partly by telomere erosion. The presence of telomerase can be considered to be a stem cell marker, although it is also found in permanent tissue culture lines, early embryos and most cancers.
In human or animal tissues, various markers have been advanced as characteristic of all stem cells. For example the cell surface glycoprotein CD34 is found on human hematopoietic stem cells (HSCs) and can be used to enrich them from bone marrow by fluorescence‐activated cell sorting (FACS). However it is also found on other cell types, such as capillary endothelial cells, and it is unclear whether it is actually necessary for the stem cell behavior of the HSC. In fact, since it is not found on mouse HSC, which are generally similar in behavior to human HSC, it is probable that it is not necessary. CD34 is not found on human embryonic stem cells or on most epithelial stem cell types, indicating that it is not a generic stem cell marker. A molecular marker which is known to be required for stem cell function is LGR5. This is an accessory receptor for the Wnt family of signaling molecules (see Chapter 7) and is found on stem cells in the intestine, hair follicle, mammary gland and stomach. These types of stem cell all depend on Wnt signaling from their environment for continued cell division, so the presence of the LGR5 is really necessary. However it is not found on other types of stem cell, so is also not a universal marker.
An interesting type of marker is that offered by dye exclusion, in particular exclusion of the Hoechst 33342 dye. This is a bisbenzimide dye, excited by UV light to emit a blue fluorescence. It is widely used as a DNA‐binding reagent, but it is also actively pumped out of some cell types. If a subgroup of cells has lost more dye than the rest of the population, then it appears in flow cytometry as a cluster of cells showing less blue fluorescence than average. This is called a side population. The side population is enriched for stem cells in some situations, especially in murine bone marrow where it provides a similar degree of enrichment of hematopoietic stem cells to FACS using a panel of cell surface markers (Figure 1.2). The dye exclusion property is due to the activity of cell membrane transporter molecules including the P‐glycoprotein (MDR1) and transporters of the ABC class. Dye exclusion is indicative of an increased capacity for export of all hydrophobic small molecules, many of which are toxic to cells. Although useful to the investigator, it is unlikel...