Nel 1859 L'origine delle specie di Charles Darwin scosse il mondo dalle fondamenta. Darwin sapeva benissimo che la sua teoria dell'evoluzione avrebbe provocato un terremoto, ma non avrebbe mai potuto immaginare che, un secolo e mezzo dopo, la controversia avrebbe continuato a infuriare. L'evoluzione è considerata un «fatto» da tutti gli scienziati autorevoli, e per la verità anche dai teologi più illuminati, eppure milioni di persone continuano a negarla o per ignoranza o per obbedienza a una religione, con risultati inquietanti.
Richard Dawkins si inserisce nel dibattito in corso e fornisce un'esauriente panoramica delle prove scientifiche dell'evoluzionismo, prendendo in esame le varie discipline, dalla chimica alla biologia, dall'embriologia alla paleontologia, e le moderne strumentazioni che contribuiscono a confermare sotto molteplici profili la realtà dell'evoluzione e, dopo aver sfatato la leggenda degli anelli mancanti (in realtà ne mancano sempre meno...), conduce il lettore lungo l'affascinante itinerario di studi aperto da Darwin.
E lo fa scavando in una miniera di evidenze scientifiche: analizza gli esempi viventi di selezione naturale e i reperti fossili, gli orologi naturali che hanno segnato le tappe del lungo processo evolutivo e le complesse fasi di sviluppo dell'embrione, le dinamiche della tettonica a placche e i meccanismi della genetica molecolare. Per giungere a una conclusione incontrovertibile: «Non è un caso che ci ritroviamo appollaiati su un ramoscello sottile in mezzo al rigoglioso, fiorente albero della vita. Non è un caso, bensì la diretta conseguenza dell'evoluzione per selezione naturale non casuale».
Mentre in tutto il mondo è in atto il tentativo di minare alla radice l'autorevolezza della scienza, con Il più grande spettacolo della Terra Dawkins sceglie di partecipare alla battaglia contro ogni vecchia e nuova forma di oscurantismo riproponendo la lucida visione delle leggi della vita che porta il nome di uno dei più grandi scienziati di ogni tempo.

eBook - ePub
Il più grande spettacolo della Terra
Perché Darwin aveva ragione
- 408 pagine
- Italian
- ePUB (disponibile su mobile)
- Disponibile su iOS e Android
eBook - ePub
Informazioni su questo libro
Scelto da 375,005 studenti
Accesso a oltre 1,5 milioni di titoli a un prezzo mensile trasparente.
Studia in modo più efficiente con i nostri strumenti dedicati.
Informazioni
Argomento
Scienze biologicheCategoria
Scienza generaleX
L’albero della cuginanza
Un osso connesso all’altro
Che capolavoro è lo scheletro dei mammiferi. Non voglio dire che sia bello, anche se penso che lo sia; voglio dire che è fantastico poter dire «lo scheletro dei mammiferi», è fantastico che una struttura così complessa sia da un lato così straordinariamente diversa, nei particolari, da un mammifero all’altro, e dall’altro chiaramente la stessa in tutti. Lo scheletro umano è talmente noto che non occorre riprodurne l’immagine, ma provate a guardare lo scheletro di un pipistrello (fig. 10.1). Non è affascinante vedere come ogni osso abbia il suo omologo riconoscibile nello scheletro umano? Riconoscibile per l’ordine in cui ognuno è unito all’altro. Solo le proporzioni sono diverse. Le zampe anteriori del pipistrello sono enormi (rispetto alle dimensioni complessive del corpo, s’intende), ma non si può non notare la corrispondenza tra le nostre dita e le lunghe ossa delle ali (ill. 27c). La mano umana e la «mano» del pipistrello sono chiaramente due versioni della stessa cosa: nessuna persona sana di mente potrebbe negarlo. Il termine tecnico per questo tipo di somiglianza assoluta è «omologia». Le ali del pipistrello e le nostre mani fatte per afferrare le cose sono «omologhe». Le mani del nostro antenato comune, nonché il resto dello scheletro, furono prese, tirate o compresse, pezzo per pezzo, in direzioni e quantità diverse lungo linee evolutive diverse.
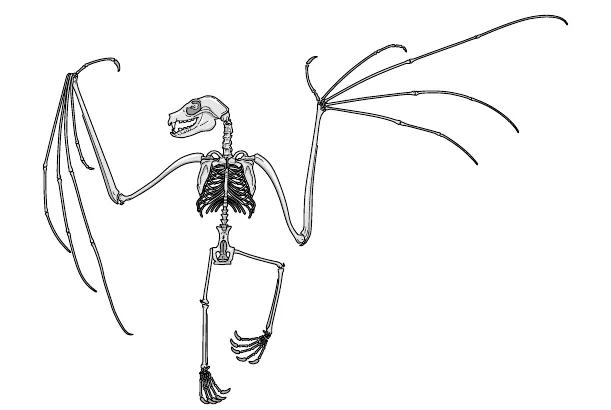
Figura 10.1. Scheletro di pipistrello.
Lo stesso vale, benché sempre in proporzioni diverse, per l’ala dello pterodattilo (fig. 10.2). Non era un mammifero, ma il principio è lo stesso, il che rende la somiglianza ancora più impressionante. La membrana alare dello pterodattilo è in gran parte sorretta da un unico dito, quello che chiameremmo «mignolo». Confesso di essere colto da nevrosi omologica al pensiero che un simile ingente peso sia trasportato dal mignolo, giacché negli esseri umani questo è il dito più piccolo e fragile. È una nevrosi sciocca, naturalmente, perché, lungi dall’essere «piccolo», il mignolo dello pterodattilo era lungo quasi quanto tutto il corpo dell’animale e probabilmente era forte e robusto come le braccia per noi. Il mignolo è stato modificato in maniera che sostenesse la membrana alare. I vari particolari si sono diversificati, ma il mignolo è ancora riconoscibile per via della sua relazione spaziale con le altre ossa dello scheletro. Questo montante lungo e robusto che sostiene le ali è «omologo» del nostro mignolo. In pterodattilese, «mignolo» significa «montante lungo e robusto».
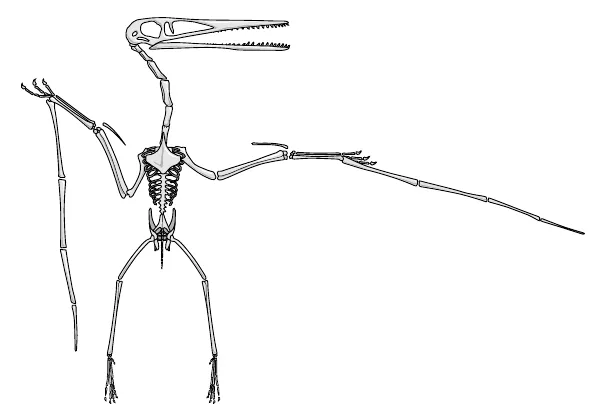
Figura 10.2. Scheletro di pterodattilo.
Oltre ai veri volatori (uccelli, pipistrelli, pterosauri e insetti), vi sono numerosi planatori, e il volo planato forse ci dice qualcosa sul volo vero e proprio. Questi animali sono dotati di patagi che hanno bisogno del supporto di uno scheletro, il quale però non è detto sia rappresentato dalle ossa delle dita, come accade nelle ali dei pipistrelli e degli pterosauri. Gli scoiattoli volanti (due gruppi indipendenti di roditori) e i petauri o «falangeri volanti» (marsupiali australiani che sono pressoché identici agli scoiattoli volanti, ma non sono loro stretti parenti) hanno una membrana che si estende dalle zampe anteriori a quelle posteriori. Le singole dita non devono sostenere un grande peso, perciò non sono grandi. Io, con la mia nevrosi del mignolo, preferirei di gran lunga essere uno scoiattolo volante che uno pterodattilo, perché mi pare «giusto» usare l’intera estensione delle «braccia» e delle «gambe» per sorreggere il peso.
Se osserviamo lo scheletro del drago volante (fig. 10.3), un altro elegante planatore della foresta, vediamo subito che nel suo caso sono state le costole, anziché le dita o le zampe anteriori e posteriori, a modificarsi per sostenere le «ali», le membrane di volo. Ancora una volta, la somiglianza con gli scheletri di altri vertebrati è evidentissima. Esaminando ogni osso uno per uno, si potrebbe trovare l’esatto corrispondente nello scheletro di un uomo o di uno pterosauro.
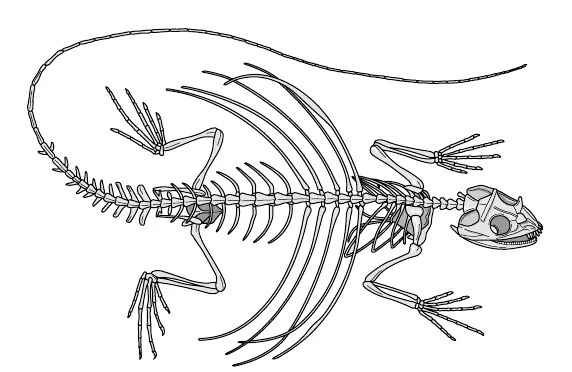
Figura 10.3. Scheletro di «drago volante».
Il colugo, o «lemure volante» (ill. 27b), delle foreste del Sudest asiatico somiglia agli scoiattoli volanti e ai petauri, solo che la coda, come le zampe anteriori e posteriori, è compresa nella struttura di sostegno del patagio. Qui non riesco a immedesimarmi, perché non posso immaginare che effetto faccia avere una coda, anche se noi esseri umani, assieme a tutti gli altri ominidi «senza coda», abbiamo un vestigio di coda (il coccige). Pressoché privi di coda come siamo, noi umani facciamo fatica a immaginare che vuol dire essere una scimmia ragno, con una coda più lunga della colonna vertebrale (ill. 26a) e delle già lunghe zampe. Come in molte platirrine del Nuovo Mondo (anzi, come generalmente in molti mammiferi del Nuovo Mondo, un fatto curioso di difficile decifrazione), la coda dell’atele, o scimmia ragno, è «prensile», ovvero si è modificata per afferrare le cose e sembra quasi terminare in una mano supplementare, benché non sia omologa di una mano vera e non abbia dita. In realtà, somiglia molto a una gamba o un braccio in più.
Lo scheletro che sorregge la coda è sostanzialmente lo stesso che sorregge la coda di qualsiasi altro mammifero, ma è modificato in maniera da svolgere un ruolo diverso. Perciò da un lato la coda dell’atele non è esattamente la stessa che negli altri mammiferi, perché ha una maggiore dotazione di vertebre, dall’altro le sue vertebre sono visibilmente le stesse di qualsiasi altra coda, compreso il nostro coccige. Vi immaginate che effetto fa essere una scimmia con cinque «mani» prensili, una all’estremità delle gambe, una all’estremità delle braccia e una coda prensile alla quale appendersi allegramente? Io no, non me lo immagino; ma so che la coda di un atele è omologa del mio coccige, proprio come il forte, lunghissimo osso alare dello pterodattilo è omologo del mio mignolo.
Veniamo a un altro dato sorprendente. Lo zoccolo del cavallo è omologo dell’unghia del dito medio delle nostre mani (o del dito medio dei nostri piedi). I cavalli camminano letteralmente in punta di piedi, o forse sarebbe meglio dire in punta del piede. Infatti hanno perso quasi del tutto le altre dita: gli omologhi del nostro indice e del nostro anulare e i loro equivalenti nelle zampe posteriori sopravvivono come piccoli «peroni» uniti al «cannone», e non sono visibili fuori della pelle. Il cannone è omologo dell’osso metacarpale umano delle nostre mani (o di quello metatarsale dei nostri piedi). L’intero peso del cavallo, che è notevole nel caso di uno Shire o un Clydesdale, si regge sui medi delle zampe anteriori e posteriori. Le omologie, per esempio con il medio dell’uomo o con quello del pipistrello, sono chiarissime. Nessuno ne può dubitare; e, quasi a voler rendere il concetto ancora più evidente, a volte nascono cavalli anomali con tre dita per zampa, nei quali il medio funge da «piede» normale, mentre le due dita che lo affiancano hanno zoccoli in miniatura (fig. 10.4).
Figura 10.4. Cavallo polidattilo.
Non è splendida l’idea di una modifica quasi indefinita che si compie nell’arco di ere geologiche e nella quale ogni forma modificata mantiene tracce inconfondibili dell’originale? Mi esaltano i litopterni, erbivori estinti del Sudamerica, che non sono strettamente imparentati con nessun animale moderno e che erano assai diversi dai cavalli, fatta eccezione per zampe e zoccoli, quasi uguali ai loro. I cavalli (in Nordamerica)* e i litopterni (in Sudamerica, che all’epoca era una gigantesca isola, giacché l’istmo di Panama era ancora di là da venire) evolsero in maniera indipendente la stessa esatta riduzione di tutte le dita eccetto il medio, e svilupparono gli stessi identici zoccoli. Forse un mammifero erbivoro non ha molti modi per diventare un veloce corridore. I cavalli e i litopterni ricorsero allo stesso sistema per accrescere la velocità: ridussero tutte le dita eccetto il medio e giunsero allo stesso risultato. Le mucche e le antilopi trovarono un’altra soluzione: ridurre le dita a due.
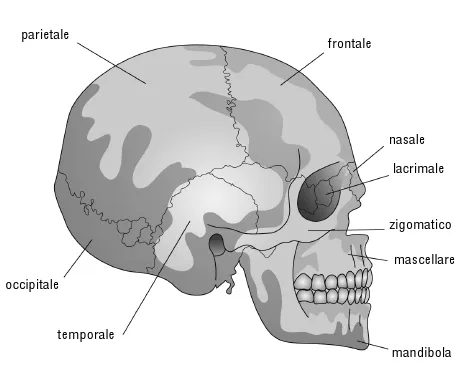
Figura 10.5. Cranio umano.
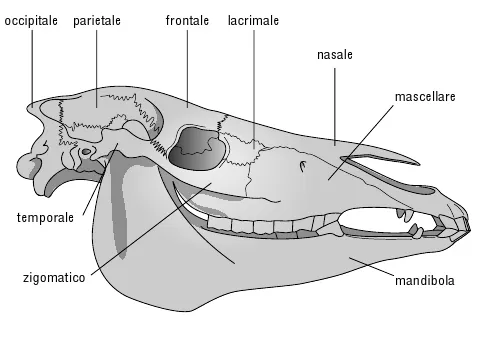
Figura 10.6. Cranio di cavallo.
L’affermazione che sto per fare potrà sembrarvi paradossale, ma capirete presto che ha senso e anzi è molto importante: gli scheletri di tutti i mammiferi sono identici, ma le singole ossa sono diverse. La chiave del paradosso sta nell’uso calcolato del termine «scheletro», inteso come sinonimo di assemblaggio di ossa connesse in maniera ordinata le une alle altre. In questo senso, la forma delle singole ossa non è affatto proprietà dello «scheletro» e lo «scheletro» ignora la forma delle singole ossa, preoccupandosi solo dell’ordine in cui esse si connettono. E ciò vale per qualsiasi mammifero, anzi per qualsiasi vertebrato terrestre. Per esempio «l’osso della testa», il cranio, contiene ventotto ossa, perlopiù connesse da rigide «suture», ma con un importante osso mobile (la mandibola).* Il bello è che, salvo qualche saltuaria eccezione, la stessa serie di ventotto ossa, etichettabili chiaramente con lo stesso nome, si rinviene in tutti i mammiferi.
La struttura è sempre la stessa, anche se la forma delle singole ossa è radicalmente diversa nei vari mammiferi.
Che conclusioni traiamo da tali osservazioni? Qui ci siamo limitati a considerare gli animali moderni, sicché non stiamo vedendo l’evoluzione in atto. Siamo i detective comparsi sulla scena dopo che il delitto è stato commesso. E lo schema delle somiglianze tra gli scheletri degli animali moderni è proprio quello che sarebbe logico aspettarsi se tutti discendessero da un antenato comune, alcuni da meno tempo di altri. Lo scheletro ancestrale si è gradualmente modificato nel corso delle ere geologiche. Alcune coppie di animali, come le giraffe e gli okapi, hanno un antenato comune recente. Non è propriamente corretto definire una giraffa un okapi che si è sviluppato in altezza, perché sono entrambi animali moderni. Ma è lecito supporre (confortati dalle prove fossili, alle quali però non accenneremo in questo capitolo in cui non si parla di fossili) che l’antenato comune somigliasse più all’okapi che alla giraffa.
Analogamente, gli impala e gli gnu sono stretti cugini, e cugini poco meno stretti delle giraffe e degli okapi. Tutti e quattro sono cugini più lontani di altri animali dal piede fesso, gli artiodattili, come i maiali e i facoceri (che sono cugini gli uni degli altri e dei pecari). Tutti gli artiodattili sono cugini più lontani di cavalli e zebre (che sono perissodattili e stretti cugini tra loro). Potremmo andare avanti così per un pezzo, raggruppando coppie di cugini in gruppi, e gruppi di gruppi di cugini e (gruppi di (gruppi di (gruppi di cugini))). Mi è venuto spontaneo usare le parentesi, e voi sapete che cosa significano. Il significato delle parentesi nella frase seguente vi sarà immediatamente chiaro, perché sapete già tutto sui primi cugini che condividono nonni, i secondi cugini che condividono bisnonni e così via.
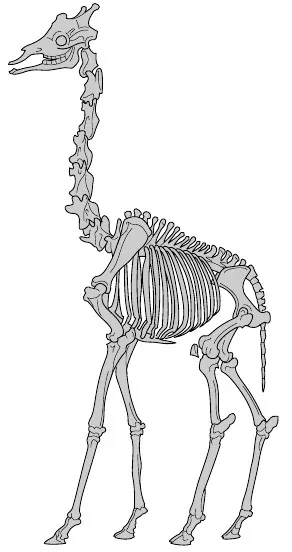 | 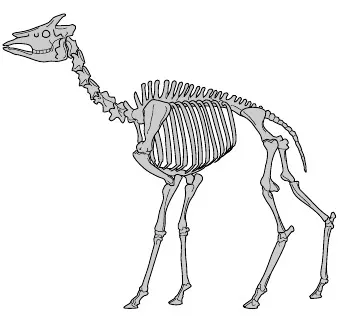 |
| Figura 10.7. Giraffa. | Figura 10.8. Okapi. |
{(lupo volpe)(leone leopardo)} {(giraffa okapi) (impala gnu)}
Tutto porta a un semplice albero ramificato degli antenati, l’albero filogenetico.
La conclusione è che l’albero delle somiglianze è in realtà un albero filogenetico. Ma ci sono interpretazioni alternative? Be’, una ce ne sarebbe... In epoca predarwiniana, i creazionisti notarono lo schema gerarchico delle somiglianze e formularono una spiegazione non evoluzionistica così poco probabile da risultare imbarazzante. A loro giudizio, le somiglianze tra esseri viventi riflettevano temi nella mente dell’artefice. L’artefice aveva varie idee su come creare animali. Ora pensava al tema dei mammiferi, ora al tema degli insetti. All’interno del tema dei mammiferi, le sue idee si bisecarono in maniera netta e gerarchica in sottotemi (per esempio il tema degli artiodattili) e in sotto-sotto-temi (per esempio il tema del maiale). L’argomentazione è fortemente speciosa, e i creazionisti oggi la citano poco. In effetti, come nel caso della distribuzione geografica di cui ho parlato nel capitolo precedente, non discutono quasi mai delle prove comparate e preferiscono incaponirsi sui fossili, un campo che hanno imparato (a torto) a ritenere promettente.
Nessun prestito
Per sottolineare quanto sia bizzarra l’immagine di un creatore rigidamente fissato con i «temi», converrà riflettere che qualsiasi progettista umano dotato di un minimo di raziocinio sarebbe ben felice di prendere un’idea da una delle sue stesse invenzioni per applicarla a un’altra invenzione. Forse nel progetto di un aereo c’è un «tema» distinto dal «tema» del progetto di un treno, ma un componente dell’aereo, per esempio un efficiente sistema di luci di cortesia sopra i sedili, potrebbe benissimo essere ricavato dal progetto dell’aereo e utilizzato sui treni. Perché questo dispositivo non dovrebbe avere un doppio uso, visto che assolverebbe in entrambi i casi la stessa funzione? All’inizio, quando furono inventate, le automobili erano chiamate «carrozze senza cavalli», il che spiega da dove fu tratta l’ispirazione. Ma i veicoli trainati da cavalli non hanno bisogno del volante, visto che la guida è data dalle redini, sicché per quel particolare componente bisognava ispirarsi a qualcos’altro. Non so a che cosa abbiano fatto riferimento gli inventori dell’auto, ma credo si siano rivolti a tutt’altro settore: quello della nautica. Prima di essere soppiantato dal volante, che fu introdotto verso la fine del XIX secolo, lo strumento originario di guida dell’auto era la barra, che, pur trovandosi nella parte anteriore del veicolo anziché in quella posteriore, si ispirava al timone delle barche.
Se le penne sono una buona idea all’interno del «tema» aviario, tanto che ne è dotato ogni uccello, volatore o inetto al volo, come mai non le ha nessun mammifero? Perché il progettista non avrebbe dovuto adottare questa ingegnosa invenzione per renderne beneficiario almeno il pipistrello? La risposta dell’evoluzionista è chiara: tutti gli uccelli hanno ereditato le penne dal loro antenato comune, che le aveva, e nessun mammifero discende da quell’antenato. Più semplice di così...* L’albero delle somiglianze è un albero filogenetico. Accade lo stesso per ogni ramo, ogni sottoramo e ogni sottosottoramo dell’albero della vita.
Veniamo ora a una questione interessante. Vi sono molti begli esempi in cui, all’apparenza, certe idee sono mutuate da una parte dell’albero e innestate su un’altra parte, come può essere innestata una varietà di mela su un determinato ceppo. Il delfino (ill. 29g), che è una piccola balena, ricorda nell’aspetto vari pesci di grandi dimensioni, fra cui la corifena cavallina (Coryphaena hippuris). Le corifene e i delfini hanno la stessa forma aerodinamica, adatta alle abitudini di vita di creature veloci che cacciano vicino alla superficie del mare. Ma nessuno dei due ha mutuato dall’altro la tecnica natatoria, che, sebbene sia all’apparenza simile, risulta diversa appena si osservano i dettagli. Benché entrambi siano veloci soprattutto grazie alla coda, la corifena, come tutti i pesci, muove la sua coda lateralmente. Il movimento laterale di flessione della spina dorsale dei pesci ancestrali è stato ereditato dalle lucertole e dai serpenti, che si potrebbe quasi dire «nuotino» sulla terraferma. Si confronti la loro andatura con il galoppo di cavalli e ghepardi. La velocità è data dalla flessione della spina dorsale, come nei pesci e nei serpenti, solo che nei mammiferi la spina dorsale si flette in su e in giù anziché di lato. Sarebbe interessante sapere in che modo abbiano compiuto questo passaggio i progenitori dei mammiferi. Forse vi fu uno stadio intermedio in cui la spina dorsale non veniva flessa quasi per niente in nessuna direzione, come accade nelle rane. I coccodrilli, invece, riescono sia a galoppare (in maniera spaventosamente veloce) sia a procedere con l’andatura da lucertola prevalente nei rett...
Indice dei contenuti
- Copertina
- Frontespizio
- Colophon
- Indice
- Prefazione
- I - Solo Una Teoria?
- II - Cani, Mucche e Cavoli
- III - Il Sentiero Fiorito Verso La Macroevoluzione
- IV - Il Silenzio e Il Tempo Lento
- V - Proprio Davanti Ai Nostri Occhi
- VI - Anello Mancante? Come Sarebbe, «Mancante»?
- VII - Ominidi Mancanti? Non Più
- VIII - Ci Sono Voluti Soltanto Nove Mesi!
- IX - L’arca Dei Continenti
- X - L’albero Della Cuginanza
- XI - La Storia Scritta Addosso
- XII - Corsa Agli Armamenti e Teodicea Evolutiva
- XIII - Vi è Qualcosa Di Grandioso in Questa Concezione Della Vita
- Appendice. I Negazionisti
- Note
- Bibliografia
- Fonti Delle Illustrazioni
- Indice Dei Nomi
- Immagini
Domande frequenti
Sì, puoi annullare l'abbonamento in qualsiasi momento dalla sezione Abbonamento nelle impostazioni del tuo account sul sito web di Perlego. L'abbonamento rimarrà attivo fino alla fine del periodo di fatturazione in corso. Scopri come annullare l'abbonamento
No, i libri non possono essere scaricati come file esterni, ad esempio in formato PDF, per essere utilizzati al di fuori di Perlego. Tuttavia, puoi scaricarli nell'app Perlego per leggerli offline su smartphone o tablet. Scopri come scaricare libri offline
Perlego offre due piani: Essential e Complete
- Essential è l'ideale per chi studia o per i professionisti che amano esplorare una vasta gamma di argomenti. Accedi all'Essential Library con oltre 800.000 titoli affidabili e bestseller nei settori business, crescita personale e discipline umanistiche. Include tempo di lettura illimitato e voce standard per la funzione Leggi ad alta voce.
- Complete: perfetto per studenti avanzati e ricercatori che necessitano di accesso completo e senza restrizioni. Sblocca oltre 1,5 milioni di libri su centinaia di argomenti, inclusi titoli accademici e specialistici. Il piano Complete offre anche funzionalità avanzate come Leggi ad alta voce Premium e Assistente alla ricerca.
Perlego è un servizio di abbonamento a libri di testo online che ti permette di accedere a un'intera biblioteca digitale a meno del prezzo di un singolo libro al mese. Con oltre 1,5 milioni di libri su più di 990 argomenti, qui trovi tutto ciò che ti serve! Scopri la nostra missione
Cerca l'icona Sintesi vocale nel prossimo libro che leggerai per verificare se è possibile riprodurre l'audio. Questo strumento permette di leggere il testo a voce alta, evidenziandolo man mano che la lettura procede. Puoi aumentare o diminuire la velocità della sintesi vocale, oppure sospendere la riproduzione. Scopri di più sulla funzione di sintesi vocale
Sì! Puoi utilizzare l'app di Perlego su dispositivi iOS e Android per leggere quando e dove vuoi, anche offline. È perfetta per gli spostamenti quotidiani o quando sei in viaggio.
I dispositivi con iOS 13 e Android 7 o versioni precedenti non sono supportati. Scopri di più su come utilizzare l'app
I dispositivi con iOS 13 e Android 7 o versioni precedenti non sono supportati. Scopri di più su come utilizzare l'app
Sì, puoi accedere a Il più grande spettacolo della Terra di Richard Dawkins, Laura Serra in formato PDF e/o ePub, così come ad altri libri molto apprezzati su Scienze biologiche e Scienza generale. Abbiamo oltre 1,5 milioni di libri disponibili nel nostro catalogo da esplorare.