Il risultato di un processo di sviluppo, ovvero l'emergere di un individuo adulto, con le sue caratteristiche e capacità specifiche, dipende da una complessa interazione fra la specificazione genetica e l'esperienza. Questo volume riunisce per la prima volta le conoscenze relative ai diversi livelli di analisi dell'argomento, dalla formazione dei circuiti neurali agli esempi di maturazione funzionale del sistema nervoso, al ruolo dell'ambiente e dell'esperienza.

- 304 pagine
- Italian
- ePUB (disponibile su mobile)
- Disponibile su iOS e Android
eBook - ePub
Informazioni su questo libro
Scelto da 375,005 studenti
Accesso a oltre 1,5 milioni di titoli a un prezzo mensile trasparente.
Studia in modo più efficiente con i nostri strumenti dedicati.
Informazioni
Argomento
PsicologiaCategoria
Psicologia dello sviluppo1. Le cellule nervose e i segnali nervosi
In questo capitolo richiamiamo sinteticamente le caratteristiche morfofunzionali delle cellule nervose, il tipo di segnali da esse generato per codificare l’informazione in ingresso e trasmetterla a distanza, e le modalità della comunicazione fra cellule nervose diverse mediante il rilascio di molecole specifiche, i neurotrasmettitori, un processo noto come trasmissione sinaptica. Dal momento che la modificabilità dell’efficacia della trasmissione sinaptica da parte dell’esperienza (plasticità sinaptica) è alla base dell’azione dell’esperienza nella guida dello sviluppo, nel box 1.1 (in fondo al capitolo) forniamo una breve presentazione dei meccanismi della plasticità sinaptica.
1.1 Il neurone
Tutti gli organi del corpo sono costituiti da cellule e il sistema nervoso non è certo un’eccezione. Questo concetto, che oggi ci appare ovvio, accese il dibattito scientifico del Diciannovesimo e dell’inizio del Ventesimo secolo. I sostenitori della teoria reticolare, di cui uno dei rappresentanti principali fu Camillo Golgi, ritenevano che il sistema nervoso fosse costituito da un intrico di fibre che non rappresentavano unità singole. I sostenitori della teoria cellulare, rappresentati principalmente da Santiago Ramón y Cajal, e che includevano anche Sigmund Freud in una sua breve incursione nel campo delle neuroscienze, sostenevano invece che il tessuto nervoso è formato da cellule. L’utilizzo di microscopi a sempre maggiore risoluzione e di opportune tecniche di colorazione delle cellule nervose, quali il metodo di Golgi, ha risolto il dilemma mostrando che gli elementi costitutivi del sistema nervoso sono racchiusi entro membrane cellulari e sono quindi cellule. In particolare, il tessuto nervoso è formato da due tipi cellulari: i neuroni e le cellule gliali. Questi due tipi di cellule svolgono ruoli diversi: ai neuroni spetta il compito di trasmettere ed elaborare l’informazione, codificata in segnali nervosi, mentre le cellule gliali facilitano la trasmissione dei segnali nervosi nei neuroni, mantengono la corretta composizione chimica del mezzo extracellulare e contribuiscono a fornire un adeguato apporto energetico alle aree cerebrali più attive, regolando il flusso sanguigno attraverso le arteriole. Recenti osservazioni indicano che gli astrociti, un particolare tipo di cellula gliale, possono anche essere coinvolti nella trasmissione dei segnali nervosi sia in ricezione, essendo in grado di rispondere ai neurotrasmettitori rilasciati dai neuroni, sia rilasciando a loro volta neurotrasmettitori capaci di attivare i neuroni. Il ruolo funzionale di questa interazione neuroni-astrociti è ancora sotto intensa analisi (Fellin et al., 2004).
I neuroni possono essere molto diversi tra loro dal punto di vista morfologico ma possiedono un’organizzazione generale comune (fig. 1.1). Tutti i neuroni hanno un corpo cellulare (detto anche soma) contenente nel nucleo il materiale genetico (DNA). Nel nucleo avvengono i processi di trascrizione, attraverso i quali il DNA viene trascritto in RNA messaggero (mRNA), che può essere in seguito tradotto in proteine (sintesi proteica) ad opera dei ribosomi. La sintesi proteica può avvenire sia a livello del corpo cellulare sia a livello degli assoni e dei dendriti.
Dal corpo cellulare emergono due tipi di ramificazioni, i dendriti e l’assone. Questi prolungamenti rappresentano la caratteristica morfologica distintiva dei neuroni. Ogni neurone possiede molti dendriti e un solo assone. Tipi neuronali diversi possono avere alberi dendritici molto diversi (fig. 1.1), ma la funzione dei dendriti rimane la stessa in tutti i neuroni. Anche la grandezza e la lunghezza degli assoni cambia fra tipi neuronali diversi, ma, anche per l’assone, la funzione rimane la stessa in tutti i neuroni.
I dendriti, insieme al soma, costituiscono il compartimento di ingresso del neurone. Infatti è sui dendriti, e in minor misura sul soma, che arrivano i segnali provenienti dagli altri neuroni. La morfologia dell’albero dendritico è una delle caratteristiche che maggiormente differenzia i diversi tipi neuronali (fig. 1.1).
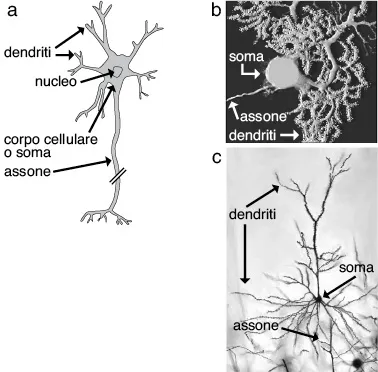
Fig. 1.1. a: Rappresentazione schematica di un tipico neurone e dei suoi diversi distretti; b e c: Esempi della morfologia di tipi neuronali diversi. In b è rappresentata una cellula del Purkinje, presente nel cervelletto; in c una cellula piramidale, presente nella corteccia cerebrale. Notare la presenza dei tipici distretti neuronali (soma, dendriti, assone) nonostante la differente morfologia che essi assumono.
L’assone conduce i segnali nervosi anche a lunga distanza e, ramificandosi, può inviare i segnali a una o più cellule bersaglio. I segnali quindi fluiscono dai dendriti, dove vengono recepiti, al corpo cellulare e all’assone, che li conduce fino al terminale assonico, dal quale vengono trasmessi alle cellule bersaglio. La trasmissione dell’informazione alla cellula bersaglio avviene attraverso specializzazioni anatomo-funzionali dei terminali assonici dette sinapsi. Per allargare la superficie di membrana disponibile alla ricezione degli ingressi sinaptici, sui dendriti di molti neuroni vi sono piccole protuberanze dette spine dendritiche. I dendriti rappresentano quindi il dominio di ricezione del neurone, l’assone rappresenta il dominio di conduzione e il terminale assonico, detto anche terminale presinaptico, rappresenta il dominio di trasmissione.
1.2 I segnali nervosi
1.2.1 Il potenziale di riposo
I segnali nervosi sono segnali elettrici. Essi consistono in variazioni del potenziale di membrana presente nelle cellule nervose a riposo, ovvero nelle cellule nervose non impegnate a ricevere e trasmettere segnali.
Se si pone un elettrodo all’interno di un neurone a riposo e si misura il potenziale elettrico, si osserva che l’interno del neurone ha un potenziale elettrico negativo rispetto all’esterno. Tale negatività interna, che può variare tra –40 mV e –90 mV (1 mV corrisponde a un millesimo di Volt) in tipi neuronali diversi, assume tipicamente il valore di –65 mV. Questa differenza di potenziale è chiamata potenziale di membrana, Vm, a riposo, in quanto è presente nel neurone inattivo. L’esistenza di un Vm a riposo deriva da un’ineguale distribuzione degli ioni ai capi della membrana che delimita il neurone (fig. 1.2). Essendo costituita da lipidi, la membrana rappresenta un ostacolo insuperabile per le molecole elettricamente cariche, che possono tuttavia attraversarla grazie a proteine specializzate dette canali ionici. Questi formano nella membrana dei pori in grado di consentire il passaggio degli ioni e sono in molti casi selettivi per il passaggio di una singola specie ionica.
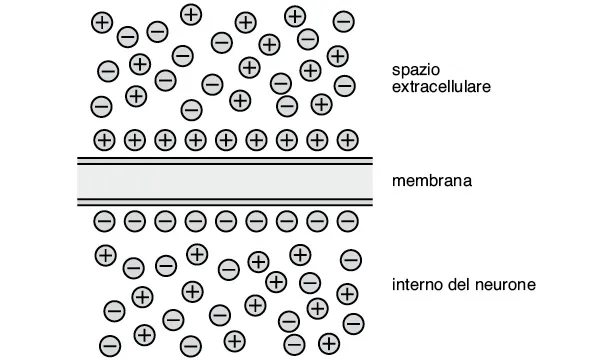
Fig. 1.2. L’esistenza di un potenziale di membrana si deve alla separazione di cariche positive e negative ai capi della membrana cellulare. L’eccesso di cariche positive all’esterno e di cariche negative all’interno costituisce una frazione molto piccola del numero totale di ioni all’esterno e all’interno del neurone. Le cariche in eccesso si posizionano immediatamente al di sotto e al di sopra della superficie interna ed esterna della membrana.
All’interno del neurone si trovano grandi molecole organiche che sono intrappolate all’interno del neurone e che hanno carica negativa. Gli ioni positivi, detti cationi, presenti nei liquidi extracellulari, subiscono quindi un’attrazione elettrostatica che li spinge a fluire verso l’interno carico negativamente. Ma non è solo l’attrazione elettrostatica che guida il fluire degli ioni attraverso la membrana: gli ioni, come le altre molecole in soluzione, tendono a diffondere da una zona a maggiore concentrazione verso una a concentrazione minore. La spinta a muoversi in base al gradiente di concentrazione si combina con la spinta fornita dalla diversa ripartizione di cariche (gradiente elettrico) nel determinare il flusso netto di uno ione fra interno ed esterno del neurone. Ad esempio, nel caso del catione potassio (K+), che è più concentrato all’interno del neurone, si avrà una spinta a muoversi attraverso la membrana in senso interno-esterno in base al gradiente di concentrazione, e una spinta in senso opposto in base al gradiente elettrico.
Una condizione stabile, in cui il flusso di potassio in entrata bilancia il flusso in uscita, si avrà quando la forza dovuta al gradiente elettrico si bilancia con quella dovuta al gradiente di concentrazione. Questo equilibrio, che si raggiunge separatamente per ciascun tipo di ione capace di attraversare la membrana, corrisponde a un Vm chiamato potenziale di equilibrio per il potassio (EK). Quando Vm è uguale a EK il neurone è in una situazione in cui il flusso di potassio netto è zero, per cui il Vm rimane costante e le concentrazioni di questo ione all’interno e all’esterno della cellula rimangono anch’esse costanti.
La relazione quantitativa fra il potenziale di equilibrio per uno ione e le concentrazioni interne ed esterne di questo stesso ione fu derivata dal chimico-fisico tedesco Walther Nernst ed è, per un catione X+, la seguente:
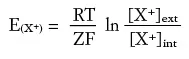
dove [X+]ext e [X+]int sono le concentrazioni interne ed esterne dello ione ed EX+ è il potenziale di equilibrio per quello ione. EX+ è proporzionale al logaritmo del rapporto fra le concentrazioni, e la proporzionalità è data dal rapporto di alcune costanti: R (costante dei gas), Z (valenza dello ione X+), F (costante di Faraday, che fornisce la carica equivalente di una mole di ioni monovalenti), T (temperatura in gradi assoluti). Una situazione in cui la concentrazione di cationi è maggiore all’esterno sarà in equilibrio per un potenziale di membrana positivo; viceversa una maggiore concentrazione di cationi all’interno della cellula sarà in equilibrio per un potenziale di membrana negativo. È importante notare che, se Vm è diverso dal potenziale di equilibrio per uno ione, ciò significa che il flusso in entrata e quello in uscita per quello ione non sono bilanciati, per cui vi sarà una corrente netta entrante o uscente. Se, per esempio, Vm è più negativo del potenziale di equilibrio per un catione, i cationi, carichi positivamente, fluiranno verso l’interno del neurone dove contribuiranno a far diventare Vm più positivo. Un flusso in direzione opposta avrà luogo se invece Vm fosse più positivo del potenziale di equilibrio. In entrambi i casi, il flusso di cationi attraverso la membrana persisterà sino a quando Vm non arriverà ad eguagliare il potenziale di equilibrio. La differenza tra Vm e il potenziale di equilibrio E(X+) è quindi la forza elettromotrice (f.e.m.) che spinge le correnti ioniche di X+ ad attraversare la membrana, come formalizzato nell’equazione
f.e.m. = Vm – E(X+)
Quali sono i cationi coinvolti nel determinare la presenza di un potenziale di riposo nelle cellule nervose? Nei neuroni esistono canali ionici sempre aperti, i cosiddetti canali passivi, che però hanno permeabilità selettiva per determinati cationi. In particolare, i canali passivi hanno un’alta permeabilità allo ione potassio e una piccola permeabilità (circa 250 volte inferiore) allo ione sodio (Na+); a riposo la permeabilità per gli altri cationi è trascurabile. Saranno quindi il potassio, e in misura minore il sodio, ad essere coinvolti nei meccanismi che generano Vm a riposo del neurone. Il potassio è circa 20 volte più concentrato all’interno che all’esterno della cellula e, utilizzando l’equazione di Nernst, possiamo calcolare che il potassio si trova in equilibrio quando Vm assume un valore di circa –75 mV. Al contrario, applicando l’equazione di Nernst al caso del sodio troveremo un potenziale di equilibrio di +55mV. Da queste considerazioni è evidente che in un neurone a riposo, che come abbiamo detto ha potenziale di membrana intorno ai –65 mV, né il potassio né il sodio sono in condizioni di equilibrio; infatti –65 mV non corrisponde al potenziale di equilibrio previsto dall’equazione di Nernst per nessuno dei due ioni. In particolare, il potassio è spinto a uscire dal neurone, dato che il potenziale di membrana è più positivo del suo potenziale di equilibrio, e il sodio ha una forte spinta a entrare. Quindi sia sodio che potassio fluiscono attraverso la membrana di un neurone a riposo, il potassio verso l’esterno e il sodio verso l’interno. La stabilità del potenziale di riposo è garantita dal fatto che vi è un perfetto bilancio tra il flusso di sodio in entrata e il flusso di potassio in uscita. In altre parole, la corrente totale (o corrente netta) che attraversa la membrana è zero e quindi non vi saranno variazioni di Vm.
Il potenziale di riposo del neurone è molto più vicino al potenziale di equilibrio del potassio (–75 mV) che non a quello del sodio (+55mV). Questo è dovuto al fatto che a determinare l’entità del flusso di una specie ionica attraverso la membrana non concorre unicamente la f.e.m. ma è importante anche quanto la membrana offre resistenza al passaggio di quel determinato ione, ovvero da quanti canali ionici permeabili a quello ione sono aperti. A parità di f.e.m. il flusso sarà maggiore se la resistenza è minore. Il potenziale di riposo risulterà più vicino al potenziale di equilibrio della specie per la quale la resistenza della membrana è minore. Come già detto, la membrana a riposo è molto più permeabile al potassio che non al sodio. Il potenziale di membrana a riposo risulterà quindi più vicino al potenziale di equilibrio del potassio. È anche spiegabile come il flusso di sodio e quello di potassio si bilancino proprio a –65mV; infatti a questo valore di potenziale di membrana il sodio ha una grande f.e.m. che lo spinge a entrare; tuttavia, il flusso sarà limitato dall’alta resistenza della membrana e si bilancerà con il flusso di potassio determinato da una tendenza a uscire meno forte di quella del sodio, ma con resistenza nettamente minore.
Il potenziale di riposo corrisponde a una situazione di equilibrio tra il flusso di sodio in entrata e quello di potassio in uscita. Si potrebbe obiettare che questa condizione è impossibile da mantenere nel tempo, in quanto la continua entrata di sodio e la continua uscita di potassio dovrebbero lentamente cambiare e infine annullare le differenze di concentrazione tra interno ed esterno. L’annullamento delle differenze di concentrazione annullerebbe praticamente il potenziale di membrana. Tuttavia questo disastroso fenomeno non avviene, in quanto i neuroni sono provvisti di un enzima, la pompa sodio-potassio ATPasi, che è posto sulla membrana e, proprio come una pompa, estrude dalla cellula 3 ioni Na+ in cambio di 2 ioni K+ in entrata (fig. 1.3). Poiché i flussi ionici prodotti dalla pompa sono elettricamente sbilanciati, la sua azione non ...
Indice dei contenuti
- Introduzione
- Parte prima. Sviluppo del sistema nervoso
- 1. Le cellule nervose e i segnali nervosi
- 2. Formazione del tubo neurale e sua suddivisione in regioni
- 3. Neurogenesi, differenziamento e sopravvivenza dei neuroni
- 4. Formazione dei circuiti neurali: allungamento degli assoni, sinaptogenesi, mielinizzazione
- Parte seconda. Sviluppo delle funzioni sensoriali e cognitive in relazione all’attività del sistema nervoso
- 5. Metodologie utilizzate in psicobiologia dello sviluppo
- 6. Funzioni sensoriali e cognitive nel feto
- 7. Lo sviluppo della funzione visiva
- 8. Lo sviluppo della funzione acustica
- 9. Lo sviluppo delle capacità mnestiche
- 10. Lo sviluppo del linguaggio
- 11. Lo sviluppo del riconoscimento dei volti
- Parte terza. Ruolo dell’esperienza e periodi critici nello sviluppo
- 12. Ruolo dell’esperienza sensoriale e periodi critici nello sviluppo del sistema visivo
- 13. Ruolo dell’esperienza sensoriale e periodi critici nello sviluppo del sistema acustico
- 14. Ruolo dell’esperienza e periodi critici nello sviluppo del linguaggio
- 15. Ruolo dell’esperienza e periodi critici nello sviluppo del riconoscimento dei volti
- 16. Plasticità intermodale durante lo sviluppo
- 17. Effetti a breve e a lungo termine delle cure parentali sullo sviluppo
- 18. Ambiente arricchito: effetti sullo sviluppo
- Bibliografia
Domande frequenti
Sì, puoi annullare l'abbonamento in qualsiasi momento dalla sezione Abbonamento nelle impostazioni del tuo account sul sito web di Perlego. L'abbonamento rimarrà attivo fino alla fine del periodo di fatturazione in corso. Scopri come annullare l'abbonamento
No, i libri non possono essere scaricati come file esterni, ad esempio in formato PDF, per essere utilizzati al di fuori di Perlego. Tuttavia, puoi scaricarli nell'app Perlego per leggerli offline su smartphone o tablet. Scopri come scaricare libri offline
Perlego offre due piani: Essential e Complete
- Essential è l'ideale per chi studia o per i professionisti che amano esplorare una vasta gamma di argomenti. Accedi all'Essential Library con oltre 800.000 titoli affidabili e bestseller nei settori business, crescita personale e discipline umanistiche. Include tempo di lettura illimitato e voce standard per la funzione Leggi ad alta voce.
- Complete: perfetto per studenti avanzati e ricercatori che necessitano di accesso completo e senza restrizioni. Sblocca oltre 1,5 milioni di libri su centinaia di argomenti, inclusi titoli accademici e specialistici. Il piano Complete offre anche funzionalità avanzate come Leggi ad alta voce Premium e Assistente alla ricerca.
Perlego è un servizio di abbonamento a libri di testo online che ti permette di accedere a un'intera biblioteca digitale a meno del prezzo di un singolo libro al mese. Con oltre 1,5 milioni di libri su più di 990 argomenti, qui trovi tutto ciò che ti serve! Scopri la nostra missione
Cerca l'icona Sintesi vocale nel prossimo libro che leggerai per verificare se è possibile riprodurre l'audio. Questo strumento permette di leggere il testo a voce alta, evidenziandolo man mano che la lettura procede. Puoi aumentare o diminuire la velocità della sintesi vocale, oppure sospendere la riproduzione. Scopri di più sulla funzione di sintesi vocale
Sì! Puoi utilizzare l'app di Perlego su dispositivi iOS e Android per leggere quando e dove vuoi, anche offline. È perfetta per gli spostamenti quotidiani o quando sei in viaggio.
I dispositivi con iOS 13 e Android 7 o versioni precedenti non sono supportati. Scopri di più su come utilizzare l'app
I dispositivi con iOS 13 e Android 7 o versioni precedenti non sono supportati. Scopri di più su come utilizzare l'app
Sì, puoi accedere a Psicobiologia dello sviluppo di Nicoletta Berardi,Tommaso Pizzorusso in formato PDF e/o ePub, così come ad altri libri molto apprezzati su Psicologia e Psicologia dello sviluppo. Abbiamo oltre 1,5 milioni di libri disponibili nel nostro catalogo da esplorare.