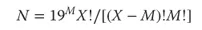Authored by one of the world's leading organic chemists, this authoritative reference provides an overview of basic strategies in directed evolution and introduces common gene mutagenesis, screening and selection methods. Throughout the text, emphasis is placed on methodology development to maximize efficiency, reliability and speed of the experiments and to provide guidelines for efficient protein engineering. Professor Reetz highlights the application of directed evolution experiments to address limitations in the field of enzyme selectivity, substrate scope, activity and robustness. He critically reviews recent developments and case studies, takes a look at future applications in the field of organic synthesis, and concludes with lessons learned from previous experiments.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
1.1 General Definition and Purpose of Directed Evolution of Enzymes
Enzymes have been used as catalysts in organic chemistry for more than a century [1a], but the general use of biocatalysis in academia and, particularly, in industry has suffered from the following often encountered limitations [1b–d]:
Limited substrate scope
Insufficient activity
Insufficient or wrong stereoselectivity
Insufficient or wrong regioselectivity
Insufficient robustness under operating conditions.
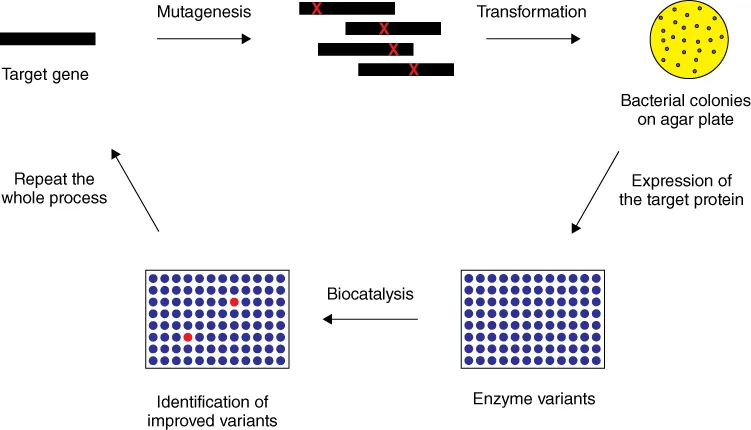
Sometimes, product inhibition also limits the use of enzymes. All of these problems can be addressed and generally solved by applying directed evolution (or laboratory evolution as it is sometimes called) [2]. It mimics Darwinian evolution as it occurs in Nature, but it does not constitute real natural evolution. The process consists of several steps, beginning with mutagenesis of the gene encoding the enzyme of interest. The library of mutated genes is then inserted into a bacterial or yeast host such as Escherichia coli or Pichia pastoris, respectively, which is plated out on agar plates. After a growth period, single colonies appear, each originating from a single cell, which now begin to express the respective protein variants. Multiple copies of transformants as well as wild-type (WT) appear, which unfortunately decrease the quality of libraries and increase the screening effort. Colony harvesting must be performed carefully, because cross-contamination leads to the formation of inseparable mixtures of mutants with concomitant misinterpretations. The colonies are picked by a robotic colony picker (or manually using toothpicks), and placed individually in the wells of 96- or 384-format microtiter plates that contain nutrient broth. Portions of each well-content are then placed in the respective wells of another microtiter plate where the screening for a given catalytic property ensues. In some (fortunate) cases, an improved variant (hit) is identified in such an initial library, which fulfills all the requirements for practical application as defined by the experimenter. If this does not happen, which generally proves to be the case, then the gene of the best variant is extracted and used as a template in the next cycle of mutagenesis/expression/screening (Scheme 1.1). This mimics “evolutionary pressure,” which is the heart of directed evolution.
Scheme 1.1 The basic steps in directed evolution of enzymes. The rectangles represent 96 well microtiter plates that contain enzyme variants, the red dots symbolizing hits.
In most directed evolution studies further cycles are necessary for obtaining the optimal catalyst, each time relying on the Darwinian character of the overall process. A crucial feature necessary for successful directed evolution is the linkage between phenotype and genotype. If a library in a recursive mode fails to harbor an improved mutant/variant, the Darwinian process ends abruptly in a local minimum on the fitness landscape. Fortunately, researchers have developed ways to escape from such local minima (“dead ends”) (see Section 4.3).
Directed evolution is thus an alternative to so-called “rational design” in which the researcher utilizes structural, mechanistic, and sequence information, possibly flanked by computational aids, in order to perform site-directed mutagenesis at a given position in a protein [3]. The molecular biological technique of site-specific mutagenesis with exchange of an amino acid at a specific position in a protein by one of the other 19 canonical amino acids was established by Michael Smith in the late 1970s [4a] which led to the Nobel Prize [4b]. The method is based on designed synthetic oligonucleotides and has been used extensively by Fersht [4c] as well as numerous other researchers in the study of enzyme mechanisms [4b]. This approach to protein engineering has also been fairly successful in thermostabilization experiments in which, for example, mutations leading to stabilizing disulfide bridges or intramolecular H-bridges are introduced “rationally” [5]. Nevertheless, in a vast number of other cases, directed evolution of protein robustness constitutes the superior strategy [6]. Moreover, when aiming for enhanced or reversed enantioselectivity, diastereoselectivity, and/or regioselectivity, rational design is much more difficult [3], in which case directed evolution is generally the preferred strategy [7]. In some cases, researchers engaging in rational design actually prepare a set of mutants, test such a “library” and even combine the designed mutations, a process that resembles “real” laboratory evolution, as shown by Bornscheuer and coworkers who generated 28 rationally designed variants of a lipase, one of them showing an improved catalytic profile [8]. Other examples are listed in Table 5.1 in Chapter 5. However, this technique has limitations, and standard directed evolution approaches are more general and most reliable.
Directed evolution of enzymes is not as straightforward as it may appear to be at this point. The challenge in putting the above principles into practice has to do with the vastness of protein sequence space. High structural diversity is easily designed in mutagenesis, but the experimenter is quickly confronted by the so-called “numbers problem” which in turn relates to the screening effort (bottleneck). When mutagenizing a given protein, the theoretical number of variants N is described by Eq. (1.1), which is based on the use of all 20 canonical amino acids as building blocks [2]:
1.1
where M denotes the total number of amino acid substitutions per enzyme molecule and X is the total number of residues (size of protein in terms of amino acids). For example, when considering an enzyme composed of 300 amino acids, 5700 different mutants are possible if one amino acid is exchanged randomly, 16 million if two substitutions occur simultaneously, and about 30 billion if three amino acids are substituted simultaneously [2].
Such calculations pinpoint a dilemma that accompanies directed evolution to this day, namely how to probe the astronomically large protein sequence space efficiently. One strategy is to limit diversity to a point at which screening can be handled within a reasonable time, but excessive diversity reduction should be avoided because then the frequency of hits in a library diminishes and may tend toward zero in extreme cases. Finding the optimal compromise constitutes the primary issue of this monograph. A very different strategy is to develop selection systems rather than experimental platforms that require screening. In a selection system, the host organism thrives and survives because it expresses a variant having the catalytic characteristics that the researcher wants to evolve. A third approach is based on the use of various types of display systems, which are sometimes called “selection systems,” although they are more related to screening. These issues are delineated in Chapter 2, which serves as a guide for choosing the appropriate system. Since it is extremely difficult to develop genuine selection systems or display platforms for directed evolution of ste...
Table of contents
Cover
Title Page
Copyright
Table of Contents
Preface
Chapter 1: Introduction to Directed Evolution
Chapter 2: Selection versus Screening in Directed Evolution
Chapter 3: Gene Mutagenesis Methods
Chapter 4: Strategies for Applying Gene Mutagenesis Methods
Chapter 5: Selected Examples of Directed Evolution of Enzymes with Emphasis on Stereo- and Regioselectivity, Substrate Scope, and/or Activity
Chapter 6: Directed Evolution of Enzyme Robustness
Chapter 7: Directed Evolution of Promiscuity: Artificial Enzymes as Catalysts in Organic Chemistry
Chapter 8: Learning from Directed Evolution
Index
End User License Agreement
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Directed Evolution of Selective Enzymes by Manfred T. Reetz in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Industrial & Technical Chemistry. We have over 1.5 million books available in our catalogue for you to explore.