- New work on the classification of the major phyla (in particular brachiopods and molluscs) has been incorporated.
- The section on trace fossils is extensively rewritten.
- The author has taken care to involve specialists in the major groups, to ensure the taxonomy is as up-to-date and accurate as possible.

- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Invertebrate Palaeontology and Evolution
About this book
Invertebrate Palaeontology and Evolution is well established as the foremost palaeontology text at the undergraduate level. This fully revised fourth edition includes a complete update of the sections on evolution and the fossil record, and the evolution of the early metazoans.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
PART ONE
General Palaeontological Concepts

Arnioceras cf. hartmanni (Oppel), an assemblage of immature ammonites from the Lower Jurassic of Black Ven, Charmouth, England (Lower Sinemurian). These specimens were probably catastrophically buried, since the soft parts must have been in place when they died preventing sediment penetrating the chambers. Painting by Raisin Moran; original specimen in the James Mitchell Museum, University College, Galway, Ireland.
1
Principles of palaeontology
1.1 Introduction
Once upon a time …
Some 4600 million years ago the Earth came into being, probably forming from a condensing disc of particles, dust and gas, which slowly rotated round the Sun. Larger particles, or planetismals, formed from this nebular disc, and as these collided they accreted, eventually forming the planets.
Of all the nine planets in the Solar System only Earth, as far as is known, supports advanced life, though at the time of writing much interest has been generated by the discovery of organic material on Jupiter’s satellite Europa. It is, however, a striking fact that life on Earth began very early indeed, within the first 30% of the planet’s history. There are remains of simple organisms (bacteria and ‘blue-green algae’, or cyanobacteria) in rocks 3400 Ma old, so life presumably originated before then. These simple forms of life seem to have dominated the scene for the next 2000 Ma, and evolution at that time was very slow. Nevertheless, the cyanobacteria and photosynthetic bacteria were instrumental in changing the environment, for they gave off oxygen into an atmosphere that was previously devoid of it, so that animal life eventually became possible.
Only when some of the early living beings of this Earth had reached a high level of physiological and reproductive organization (and most particularly when sexual reproduction originated) was the rate of evolutionary change accelerated, and with it all manner of new possibilities were opened up to evolving life. This was not until comparatively late in geological history, and there are no fossil animals known from sediments older than about 700 Ma. Needless to say, these are all invertebrate animals lacking backbones. All of them are marine; there is no record of terrestrial animals until much later. In terms of our understanding of the history of life, perhaps the most significant of all events took place about 543 Ma ago at about the beginning of the Cambrian Period, for at this stage there was a sudden proliferation of different kinds of marine invertebrates. During this critical period the principal invertebrate groups were established, and they then diversified and expanded. Some of these organisms acquired hard shells and were capable of being fossilized, and only because of this can there be any chance of understanding the history of invertebrate life.
The stratified sedimentary rocks laid down since the early Cambrian, and built up throughout the whole of Phanerozoic time, are distinguished by a rich heritage of the fossil remains of the invertebrates that evolved through successive historical periods; their study is the domain of invertebrate palaeontology and the subject of this book.
1.2 Occurrence of invertebrate fossils in Phanerozoic rocks
Hard-part preservation
Fossil invertebrates occur in many kinds of sedimentary rock deposited in the seas during the Phanerozoic. They may be very abundant in limestones, shales, siltstones and mudstones but on the whole are not common in sandstones. Sedimentary ironstones may have rich fossil remains. Occasionally they are found in some coarse rocks such as greywackes and even conglomerates. The state of preservation of fossils varies greatly, depending on the structure and composition of the original shell, the nature and grain size of the enclosing sediment, the chemical conditions at the time of sedimentation, and the subsequent processes of diagenesis (chemical and physical changes) taking place in the rock after deposition.
The study of the processes leading to fossilization is known as taphonomy. In most cases only the hard parts of fossil animals are preserved, and for these to be fossilized, rapid burial is normally a prerequisite. The soft-bodied elements in the fauna, and those forms with thin organic shells, did not normally survive diagenesis and hence have left little or no evidence of their existence other than records of their activity in the form of trace fossils. What we can see in the rocks is therefore only a narrow band in a whole spectrum of the organisms that were once living; only very rarely have there been found beds containing some or all of the soft-bodied elements in the fauna as well. These are immensely significant for palaeontology.
The oldest such fauna is of late Precambrian age, some 615 Ma old, and is our only record of animal life before the Cambrian. Another such ‘window’ is known in Middle Cambrian rocks from British Columbia, where in addition to the normally expected trilobites and brachiopods there is a great range of soft-bodied and thin-shelled animals – sponges, worms, jellyfish, small shrimp-like creatures and animals of quite unknown affinities – which are the only trace of a diverse fauna which would otherwise be quite unknown (Chapter 12). There are similar ‘windows’ at other levels in the geological column, likewise illuminating.
The fossil record is, as a guide to the evolution of ancient life, unquestionably limited, patchy and incomplete. Usually only the hard-shelled elements in the biota (apart from trace fossils) are preserved, and the fossil assemblages present in the rock may have been transported some distance, abraded, damaged and mixed with elements of other faunas. Even if thick-shelled animals were originally present in a fauna, they may not be preserved; in sandy sediments in which the circulating waters are acidic, for instance, calcareous shells may dissolve within a few years before the sediment is compacted into rock. Since the sea floor is not always a region of continuous sediment deposition, many apparently continuous sedimentary sequences contain numerous small-scale breaks (diastems) representing periods of winnowing and erosion. Any shells on the sea floor during these erosion periods would probably be transported or destroyed – another limitation on the adequacy of the fossil record.
On the other hand, some marine invertebrates found in certain rock types have been preserved abundantly and in exquisite detail, so that it is possible to infer much about their biology from their remains. Many of the best-preserved fossils come from limestones or from silty sediments with a high calcareous content. In these (Fig. 1.1) the original calcareous shells may be retained in the fossil state with relatively little alteration, depending upon the chemical conditions within the sediment at the time of deposition and after.
A sediment often consists of components derived from various environments, and when all of these, including decaying organisms, dead shells and sedimentary particles, are thrown together the chemical balance is unstable. The sediment will be in chemical equilibrium only after diagenetic physicochemical alterations have taken place. These may involve recrystallization and the growth of new minerals (authigenesis) as well as cementation and compaction of the rock (lithification), and during any one of these processes the fossils may be altered or destroyed. Shells that are originally of calcite preserve best; aragonite is a less stable form of calcium carbonate secreted by certain living organisms (e.g. corals) and is often recrystallized to calcite during diagenesis or dissolved away completely.
Calcareous skeletons preserved in more sandy or silty sediments may dissolve after the sediment has hardened or during weathering of the rock long after its induration. Moulds (often miscalled casts) of the external and internal surfaces of the fossil may be left, and if the sediment is fine enough the details these show may be very good. Some methods for the study of such moulds are described in section 7.2, with reference to brachiopods. If a fossil encloses an originally hollow space, as for instance between the pair of shells of a bivalve or brachiopod, this space may either be left empty or become filled with sediment. In the latter case a sediment core is preserved, which comes out intact when the rock is cracked open. This bears upon its surface an internal mould of the fossil shell, whereas external moulds are left in the cavity from which it came. In rare circumstances the core or the shell, or both, may be replaced by an entirely different mineral, as happens in fossils preserved in ironstones. If the original spaces in the shell are impregnated with extra minerals, it is said to be permineralized, while the growth of secondary minerals at the expense of the shell is replacement. Cores may sometimes be of pyrite. Graptolites are often preserved like this, anaeorbic decay having released hydrogen sulphide, which reacted with ferrous (Fe2+) ions in the water to allow an internal pyrite core to form. Sometimes a core of silica is found within an unaltered calcite shell. This has happened with some of the Cretaceous sea urchins of southern England. They lived in or on a sediment of calcareous mud along with many sponges, which secreted spicules of biogenic silica as a skeleton. In alkaline conditions (above pH 9), which may sometimes be generated during bacterial decay, the solubility of the silica increases markedly, and the silica so released will travel through the rock and precipitate wherever the pH is lower. The inside of a sea urchin decaying under different conditions would trap just such an internal microenvironment, within which the silica could precipitate as a gel. Such siliceous cores retain excellent features, preserving the internal morphology of the shell. On the other hand, silica may replace calcite as a very thin shell over the surface of a fossil as a result of some complex surface reactions. These siliceous crusts may retain a very detailed expression of the surface of the fossil and, since they can be treed from the rock by dissolving the limestone with hydrochloric acid, individual small fossils preserved in this way can be studied in three dimensions. Some of the most exquisite of all trilobites and brachiopods are known from material such as this.
Figure 1.1 Possible processes of fossilization of a bivalve shell: (a) original shell, buried in mud (left) or carbonate (right); (b) the shell was calcite, was buried in a carbonate sediment and was preserved intact other than as a small crystallized patch; (c) shell originally of aragonite, now recrystallized to calcite which destroys the fine structure; (d) original calcite shell retained surrounding a diagenetic core of silica; (e) a silica rim growing on the outside of the shell; (f) tectonic distortion of a shell preserved in mudstone; (g) shell preserved in mud with original shell material leached away, leaving an external and an internal mould, surrounding a mudstone core; (h) a calcareous concretion growing round the shell and inside (if the original cavity was empty), with patches of pyrite in places; (if ironstone replacement of core and part of shell.
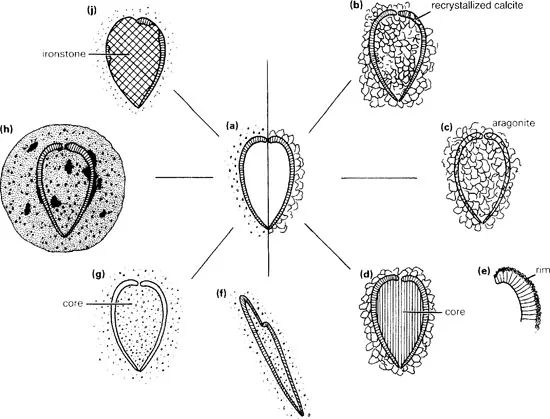
A relatively uncommon but exquisite mode of preservation is phosphatization. Sometimes the external skeleton, especially of thin organic-shelled animals, may be replaced or overgrown by a thin sheet of phosphate, or the latter may reinforce an originally phosphatic shell. In the former situation the external form of the body is precisely replicated. Alternatively a phosphatic filling of the interior of the shell may form a core, picking out internal structures in remarkable detail. Such preservation is probably associated with bacterial activity directly after the death of the animal. Many small Cambrian fossils have been preserved by phosphatization (Chapters 3 and 12), but much larger fossils may be preserved also, for example crustaceans with a fluorapatite infilling and with all their delicate appendages intact.
Fossils are often found in concretions: calcareous or siliceous masses formed around the fossil shortly after its death and burial. Concretions form under certain conditions only, where a delicate chemical balance exists between the water and sediment, by processes as yet not fully understood.
Soft-part preservation
In very rare circumstances soft-bodied organisms can be preserved as fossils, and these provide otherwise unobtainable evidence of the diversity of metazoans living at particular periods; this is discussed in Chapter 12.
1.3 Divisions of invertebrate palaeontology
Invertebrate palaeontology is normally studied as a subdivision of geology, as it is within Earth science that its greatest applications lie. It can also be seen as a biological subject, but one that has the unique perspective of geological time. Within the domain of invertebrate palaeontology there are a number of interrelated topics (Fig. 1.2), all of which have a bearing on the others and which also link up with other sciences.
Three main categories of fossils may be distinguished: (1) body fossils, in other words the actual remains of some part, usually a shell of skeleton, of a once-living organism; (2) trace fossils, which are tracks, trails, burrows or other evidence of the activity of an animal of former times – sometimes these are the only guide to the former presence of soft-bodied animals in a particular environment; (3) chemical fossils, relics of biogenic organic compounds which may be detected geochemically in the rocks.
At the heart of invertebrate palaeontology stands taxonomy; the classification of fossil and modern animals into ordered and natural groupings. These groupings, known as taxa, must be named and arranged in a hierarchial system in which their relationships are made clear, and as far as possible must be seen in evolutionary perspective.
Evolution theory is compounded of various disciplines – pure biology, comparative anatomy, embryology, genetics and population biology – but i...
Table of contents
- Cover
- Contents
- Title Page
- Copyright
- Dedication
- Preface
- PaleoBase—Macrofossils on CD-ROM
- PART ONE: General Palaeontological Concepts
- PART TWO: Invertebrate Phyla
- Systematic index
- General index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Invertebrate Palaeontology and Evolution by E. N. K. Clarkson in PDF and/or ePUB format, as well as other popular books in Naturwissenschaften & Geologie & Geowissenschaften. We have over 1.5 million books available in our catalogue for you to explore.