![]()
Part 1
Microbial physiology
![]()
1
Microbial cell structure and function
The cell is the basic unit of all living organisms, many of which are unicellular, whereas others are multicellular forms, enabling cell specialization. All cells are filled with a fluid matrix and surrounded by a cytoplasmic membrane, primarily composed of lipids and proteins. They also contain nucleic acids, the physical carriers of genetic information, along with ribosomes that take part in protein synthesis. Cells are divided into two categories: those of archaeans and eubacteria are prokaryotic, whereas the cells of fungi, protozoa, algae and other plants, and animals are eukaryotic.
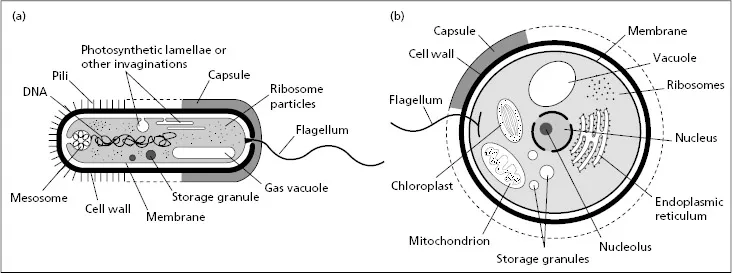
Prokaryotic cells are normally less than 5μm in diameter, with a very few notable exceptions such as the marine bacterium, Thiomargarita namibiensis, at over 100 times larger. Prokaryotes rarely possess membrane-bound organelles and have little recognizable internal ultrastructure, apart from inclusion bodies (granules of organic or inorganic compounds), various vacuoles and some specialized invaginations of the cell membrane, e.g. the lamellae of photosynthetic cells (Fig. 1.1a). Most prokaryotic cells contain a single chromosome composed of deoxyribonucleic acid (DNA), which is located in a region of the cell referred to as the nucleoid. The chromosome is usually circular, although in some prokaryotes it is linear. Prokaryotic ribosomes are 70S (Svedberg units), which refers to their rate of sedimentation on centrifugation and is a measure of their size, although density and shape of the particle can also influence this value. Almost all prokaryotes have cell walls or cell envelopes located outside the cytoplasmic membrane, which usually contain some peptidoglycan. Outside this wall they may have capsules or slime coats and propelling flagellae that are less complex than those of eukaryotic cells. Cell division in prokaryotes is normally by simple binary fission.
Eukaryotic cells are generally larger than those of prokaryotes and contain a range of membrane-bound organelles, including mitochondria, lysosomes, Golgi bodies and an extensive endoplasmic reticulum (Fig. 1.1b). Photosynthetic cells also contain chloroplasts.
The DNA of eukaryotic cells, in the form of several linear chromosomes, is characteristically complexed with histone proteins and is housed in a double membrane-bound nucleus. Eukaryotic ribosomes, apart from those located within certain organelles, are 80 S, somewhat larger than those of prokaryotes. If cell walls are present, they are composed of materials other than peptidoglycan, such as cellulose and related β-glucans, chitin or silica. Eukaryotic cells divide by a complex process of mitosis and usually have a sexual life-cycle, involving meiosis (reduction division). This process halves the number of chromosomes from diploid (2n) chromosome pairs to produce haploid (n) cells containing a single set of chromosomes, facilitates genetic recombination and results in the formation of gametes.
Prokaryotes
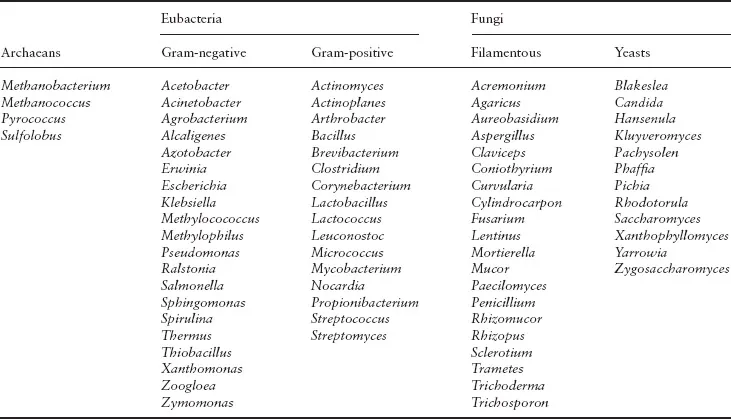
Prokaryotes have been separated into two distinct groups on the basis of the study of phylogenetic (evolutionary) relationships. They are the archaebacteria or archaea (‘ancient’ bacteria) and the eubacteria (‘true’ bacteria), the group that contains almost all established industrial prokaryotes (Table 1.1).
Archaea
These prokaryotes are quite different from eubacteria and have some features, especially aspects of the transcription and translation machinery associated with protein synthesis, that are similar to eukaryotic cells. Most archaeans live in extreme environments similar to those that early life forms are thought to have endured. Three basic physiological types are found, namely halophiles (adapted to high salt concentrations), methanogens (methane producers) and thermophiles (adapted to high temperatures), and some of these are also barophiles (adapted to high pressure) (see Chapter 2). Archaeans have relatively small genomes containing less than half the DNA of eubacteria. For example, the genome of Methanococcus jannaschii has been sequenced and found to contain 1760 genes composed of 1700 kilobase pairs (kbp). Although few archaeans are currently used for industrial purposes, they possess many interesting properties that could be exploited for biotechnological uses in the future. Their enzymes are of particular interest.
Table 1.1 Microbial genera with established industrial roles (for specific examples of their roles see Table 4.1)
Fig. 1.1 Diagrammatic representation of the main structures of (a) prokaryotic and (b) eukaryotic microbial cells. Not all structures are always present, including capsules, chloroplasts, flagellae, pili, storage granules and vacuoles (from Dawes & Sutherland (1992)).
All prokaryotes may be initially subdivided according to their characteristics in the Gram staining procedure, which is determined by cell wall/envelope structure. Archaeal cell wall composition varies considerably, some appearing Gram-positive whereas others are Gram-negative. Their cell wall constituents are quite different from those of the eubacteria, in that they contain the unique polymers, methanochondroitin and pseudomurein. They also possess distinctive membrane lipids. The archaea may be divided into three kingdoms.
1 Euryarchaeota are primarily methanogens, such as Methanobacterium and Methanosarcina, and the extreme halophiles Halobacterium and Halococcus.
2 Crenarchaeota are mostly extreme barophiles and thermophiles that include Pyrodictium, Pyrolobus, Sulfolobus and Thermoproteus.
3 Korarchaeota are hyperthermophiles which, as yet, have not been isolated as pure cultures.
Eubacteria
The eubacteria are a very diverse group that may be divided into 12 subgroups. However, almost all industrial bacteria are contained within just two of them, the proteobacteria and the Gram-positive eubacteria.
1 The Proteobacteria is a major kingdom of Gram-negative bacteria that is divided into five groups, α, β, γ, δ, and ε. They include purple photosynthetic bacteria and non-photosynthetic relatives, notably the Enterobacteriaceae (e.g. Escherichia coli), along with Hyphomicrobium, Nitrobacter, Pseudomonas, Thiobacillus and Vibrio.
2 The Gram-positive eubacteria are composed of two major subdivisions:
(a) the low guanine (G) + cytosine (C) group, which refers to the proportion of this base pair within the organism’s DNA and includes Bacillus, Clostridium, Lactobacillus, Leuconostoc, Staphylococcus, Streptococcus and Mycoplasma; and
(b) the high G + C group, which contains the actinomycetes (filamentous bacteria, e.g. Streptomyces), Corynebacterium, Mycobacterium and Micrococcus.
The other subgroups, which contain few examples of industrial bacteria, are as follows.
3 Cyanobacteria and relatives, which are oxygenic phototrophs, e.g. Anabaena, Nostoc and Spirulina.
4 Chlamydia, a group of obligate intracellular parasites.
5 Planctomyces and Pirella, bacteria lacking peptidoglycan; some with membrane-bound nucleoid.
6 Bacteroides and Flavobacteria, a subgroup that contains a mixture of physiological types.
7 Green sulphur bacteria, such as Chlorobium, an anaerobic phototroph.
8 Spirochetes and relatives which are Gram-negative coiled bacteria.
9 Deinococci, radioresistant micrococci and relatives, e.g. Deinococcus radiodurans and Thermus aquaticus.
10 Green non-sulphur bacteria and anaerobic phototrophs.
11 Thermotoga and Thermosulfobacteria, thermophiles from hot springs and marine sediments.
12 Aquiflex, a group of obligate chemolithotrophic and autotrophic hyperthermophiles.
There is probably no such thing as a typical prokaryotic cell. There is a vast amount of diversity, including: morphological diversity (size and shape; rods, cocci, spirals, filaments, etc.), structural diversity (Gram-positive or Gram-negative cell walls/envelopes, external structures such as flagellae and pili, and the ability to form spores), along with metabolic, ecological and behavioural diversity. However, it is worthwhile considering, in some detail, the key cellular features of examples of industrially valuable Gram-negative and Gram-positive bacteria.
ESCHERICHIA COLI, A GRAM-NEGATIVE BACTERIUM
E. coli was discovered in 1885 by the German bacteriologist Theodor Escherich. It is a major inhabitant of the colon of humans and the lower gut of other warmblooded animals. Some strains can cause food- and water-borne diseases that produce diarrhoea and can be especially problematical for human infants and young animals. A particularly virulent strain is E. coli 0157:H7, the cause of haemorrhagic colitis, which has recently emerged and has been associated with the ingestion of undercooked beef and raw milk.
Extensive information has been accumulated about the biochemistry, physiology and genetics of E. coli. This knowledge and the organism’s rapid growth rate, with a doubling time as low as 20 min (see Chapter 2), has led to it becoming...