![]()
Part One
Locomotion and Sensory Capabilities in Marine Fish
![]()
Chapter 1
Swimming in Marine Fish
John J. Videler and Pingguo He
1.1 INTRODUCTION
After 500 million years of natural selection, fish are extremely well adapted to various constraints set by the aquatic environment in which they live. In the dense fluid medium, they are usually neutrally buoyant and use body movements to induce reactive forces from the water to propel themselves. Animal movements are powered by contracting muscles, and these movements consume energy. The basic principles of fish locomotion are used by approximately 25,000 extant species. The variation in swimming styles, within the limits of these principles, is great. Swimming includes steady swimming at various speeds, accelerating, braking, maneuvering, jumping, diving downward, and swimming upward. Swimming behavior is different for every species and changes in each individual during growth from larva to adult. Speed, agility, and endurance maxima determine the chances for survival to a considerable extent. Peak performance in absolute terms is positively related to temperature and body length. Ultimately, performance affects the evolutionary fitness of each individual and is a significant factor that is directly related to capture by or escape from both active (e.g., trawls and seines) and passive (e.g., gillnet, longlines, and traps) fishing gear. This chapter reviews the current knowledge about fish swimming mechanisms and abilities to provide a background for discussions in later chapters.
1.1 THE SWIMMING APPARATUS
Fish are aquatic vertebrates with a skull, a vertebral column supporting a medial septum that divides the fish into two lateral halves, and lateral longitudinal muscles that are segmentally arranged in blocks, or myotomes. The vertebral column is laterally highly flexible and virtually incompressible longitudinally. Consequently, contraction of the muscles on one side of the body bends the fish, and waves of curvature along the body can be generated by series of alternating contractions on the left and the right side (Videler and Wardle 1991). Fish vertebrae are concave fore and aft (amphicoelous) and fitted with a neural arch and spine on the dorsal side. In the abdominal region, lateral projections are connected with the ribs enclosing the abdominal cavity. The vertebrae in the caudal region bear a hemal arch and spine. Neural and hemal spines point obliquely backward. The number of vertebrae varies greatly among species—European eels (Anguilla anguilla) have 114 vertebrae, and the numbers in the large order of Perciformes vary between 23 and 40 (Ford 1937). The number is not necessarily constant within a species. Atlantic herring (Clupea harengus), for example, can have between 54 and 58 vertebrae (Harder 1975a, 1975b) . The end of the vertebral column is commonly adapted to accommodate the attachment of the tail fin. Several vertebrae and their arches and spines are partly rudimentary and have changed shape to contribute to the formation of platelike structures providing support for the fin rays of the caudal fin. Most fish species have unpaired dorsal, caudal, and anal fins and paired pectoral and pelvic fins. Each fin is powered by intrinsic musculature.
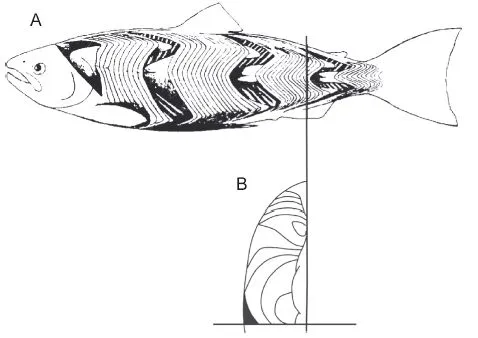
The lateral muscles are usually the main target of the fishing industry. Relatively short lateral muscle fibers are packed into myotomes between sheets of collagenous myosepts. The myotomes are cone shaped and stacked in a segmental arrangement on both sides of the median septum (Fig. 1.1 A). In cross sections through the caudal region, the muscles are arranged in four compartments. On each side is a dorsal and a ventral compartment; in some groups, they are separated by a horizontal septum. The left and right halves and the dorsal and ventral moieties are mirror images of each other. In cross sections, the myosepts are visible as more or less concentric circles of collagen. The color of the muscle fibers may be red, white, or intermediate in different locations in the myotomes (Fig. 1.1B), which was first described by Lorenzini in 1678 (Bone 1966). Red fibers are usually situated directly under the skin. The deeper white fibers form the bulk of lateral muscles, and in some species intermediately colored pink fibers are found between the two. The red fibers are slow but virtually inexhaustible and their metabolism is aerobic. They react to a single stimulus owing to the high density of nerve terminals on the fibers. The white fibers are fast, exhaust quickly, and use anaerobic metabolic pathways. White fibers are either focally or multiply innervated. Pink fibers are intermediate in most aspects. The red muscles of some large tuna and shark species are positioned well inside the white muscle mass, an arrangement that can increase the muscle temperature by as much as 10°C during swimming (Carey and Teal 1969). The final paragraph describes how this halves the twitch contraction time of the white muscles and doubles the maximum swimming speed.
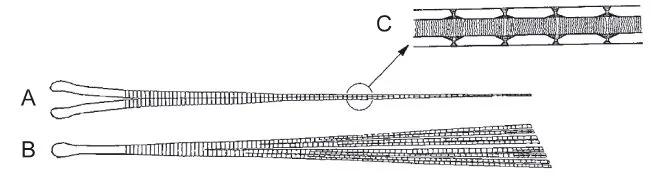
Fish fins consist of two layers of skin, usually supported by fin rays that are connected to supporting skeletal elements inside the main body of the fish. Intrinsic fin muscles find their origin usually on the supporting skeleton and insert on the fin rays. The fins of elasmobranchs (sharks and rays) are permanently extended and rather rigid compared with those of teleosts (bony fish). Elasmobranch fin rays consist of rows of longitudinally connected small pieces of cartilage in a juxtaposed arrangement. Intrinsic muscles on both sides of the rows running from the fin base to the edge bend these fins. Teleost fins can be spread, closed, and folded against the body. There are two kinds of teleost fin rays: spiny, stiff unsegmented rays and flexible segmented ones. Spiny rays stiffen the fin and are commonly used for defense. The flexible rays (Fig. 1.2) play an important role in adjusting the stiffness and camber of the fins during locomotion. They consist of mirror-)mage halves, each of which has a skeleton of bony elements interconnected by collage-nous fibers. Muscles pulling harder on the fin ray head on one side will shift the two halves with respect to each other and bend the ray or increase the stiffness against bending forces from the water. In contrast to elasmobranch fins, there are no muscles on the fin itself.
The body shape of fish may vary greatly among species, but the best pelagic swimmers have a common form. Their bodies are streamlined, with gradually increasing thickness from the point of the snout to the thickest part at about one-third of the length. From that point, the thickness gradually decreases toward the narrow caudal peduncle. A moving body in water encounters friction and pressure drag. Friction drag is proportional to the surface area, and pressure drag is proportional to the area of the largest cross section. A spherical body has the lowest friction for a given volume; a needle-shaped body encounters minimal amounts of pressure drag. An optimally streamlined body is a hybrid between a sphere and a needle and offers the smallest total drag for the largest volume. It has a diameter-to-length ratio between 0.22 and 0.24. The best pelagic swimmers have near-optimal thickness-to-length ratios (Hertel 1966).
The mechanically important part of fish skin is the tissue (the stratum compactum) underneath the scales, which consists of layers of parallel collag-enous fibers (see Videler 1993 for a review). The fibers in adjacent layers are oriented in different directions, and the angles between the layers vary between 50 and 90 degrees, but the direction in every second layer is the same. The packing of layers resembles the structure of plywood, except that in the fish stratum compactum there are also radial bundles of collagen connecting the layers; the number of layers varies between 10 and 50. In each layer, the fibers follow left-and right-handed helices over the body surface. The angle between the fibers and the longitudinal axis of the fish decreases toward the tail. In some species, the stratum compactum is firmly connected to the myo-septs in the zone occupied by the red muscle fibers; in other fish, there is no such connection. The strongest fish skins that have been tested are those of eel and shark. Values of Young’s modulus (the force per unit cross-sectional area that would be required to double the length) of up to 0.43 GPa (1GPa = 109 N/m2) have been measured. This is about one-third of the strength of mammalian tendon, for which values of 1.5 GPa have been measured.
Scales are usually found at the interface between fish and water. Several swimming-related functions have been suggested. Scales might serve to prevent transverse folds on the sides of strongly undulating fish, keeping the outer surface smooth. Spines, dents, and tubercles on scales are usually arranged to form grooves in a direction of the flow along the fish. Roughness due to microstructures on scales in general creates small-scale turbulence, which could delay or prevent the development of drag-increasing large-scale turbulence (Aleyev 1977 ).
Fish mucus covering the scales is supposed to reduce friction with the water during swimming. This assumption is based on the idea that mucus shows the “Toms effect” (Lumley 1969), which implies that small amounts of polymers are released that preclude sudden pressure drops in the passing fluid. Measurements of the effects of fish mucus on the flow show contradictory results varying from a drag reduction of almost 66% (Pacific barracuda, Sphyraena argentea) to no effect at all (Pacific bonito, Sarda chiliensis) (Rosen and Cornford 1971). Experiments with rainbow trout showed that mucus increases the thickness of the boundary layer (Daniel 1981). The layer of water around the fish is affected by the presence of the fish during swimming. Viscosity causes a layer of water close to the fish to travel with it at the speed of the fish. There is a gradient of decreasing water velocities in a direction away from the surface of the fish. The thickness of the boundary layer is defined as the distance from the surface of the fish to where the water is no longer dragged along. A thick boundary layer implies that the gradient is gradual, which reduces viscous friction. However, the penalty for a thicker boundary layer is that the fish has to drag along a larger amount of water. The conclusion might be that the effect of mucus is beneficial during slow-speed cruising but detrimental during fast swimming and acceleration (Videler 1995 ).
1.3 SWIMMING-RELATED ADAPTATIONS
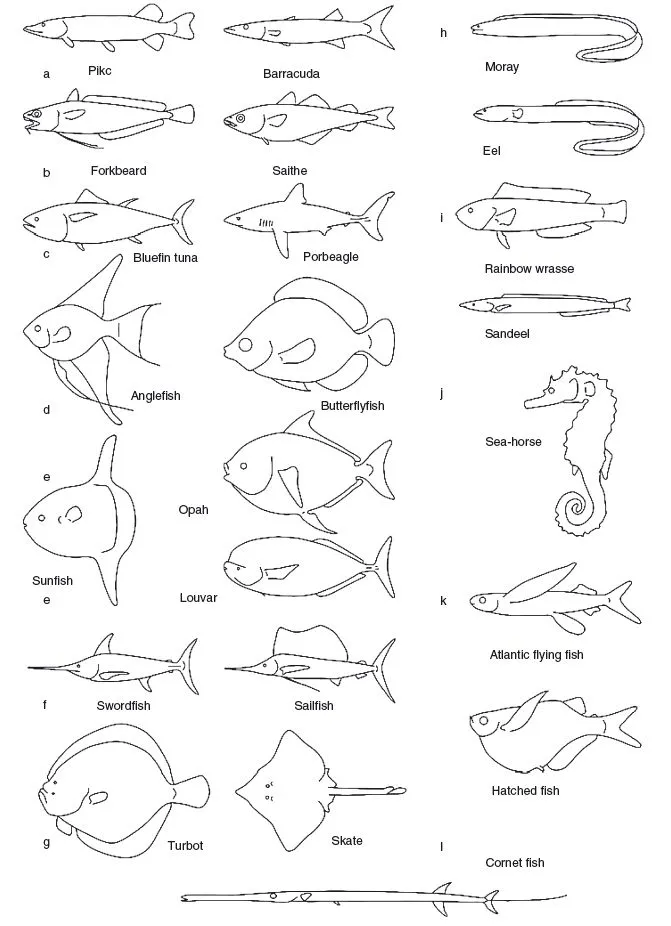
Some fish species are adapted to perform some aspect of locomotion extremely well, whereas others have a more general ability to move about and are specialized for different traits not related to swimming. Generalists can be expected to have bodies that give them moderately good performance in various special functions. Specialists perform exceptionally well in particular skills. Fast accelerating, braking, high-speed cruising, and complex maneuvering are obvious examples. The special swimming adaptations shown in Figure 1.3 are only a few of a wealth of possible examples. A closer study of the swimming habits of a large number of species will show many more specialist groups than the dozen or so described here (e.g., Lindsey 1978).
Specialists in accelerating, such as the pike (Esox lucius) and the barracuda (Fig. 1.3a), are often ambush predators. They remain stationary or swim very slowly until a potential prey is within striking distance. These species have a reasonably streamlined body and large dorsal and anal fins positioned extremely rearward, close to the caudal fin. Acceleration during the strike is achieved by the first two beats of the tail, the effect is enlarged by the rearward position of the dorsal and anal fins. The relative skin mass of the pike is reduced, compared with that of other fish, increasing the relative amount of muscles and decreasing the dead mass that has to be accelerated with the fish at each strike. Maximum acceleration rates measured for pike vary between 40 and 150 m/s2-which is 4 to 15 times the acceleration due to gravity (G = 9.8m/s2). The highest peak acceleration value reported for pike is 25 times gravity (Harper and Blake 1990, 1991).
Braking is difficult while moving in a fluid medium. Fish use the unpaired fins and tail, usually in combination with the pectoral and pelvic fins, for braking. Gadoids with multiple or long unpaired fins are good at braking. Forkbeard (Phycis phycis) swims fast and close to the bottom with elongated pelvic fins extended laterally for the detection of bottom-dwelling shrimp. The fish instantly spreads the long dorsal and anal fins and throws its body into an S-shape when a prey item is touched. In the process, the fin rays of the tail fin are actively bent forward. Braking is so effective that the shrimp has not yet reached the caudal peduncle before the fish has stopped and turned to catch it. The highest deceleration rate measured is 8.7 m/s2 for saithe (Pollachius virens) (Geerlink 1987). The contribution of the pectorals to the braking force is about 30%; the rest comes from the curved body and extended median fins (Geerlink 1987) (Fig. 1.3b).
Cruising specialists (Fig. 1.3c) migrate over long distances, swimming continuously at a fair speed. Many are found among scombrids and pelagic sharks, for example. Cruisers have highly streamlined bodies, narrow caudal peduncles with lateral keels, and high-aspect-ratio tails (aspect ratio is the tail height squared divided by tail surface area). The bluefin tuna (Thunnus thynnus), for example, is an extreme long distance swimmer, crossing the Atlantic twice a year. The body dimensions are very close to the optimum values, with a thickness--o-length ratio near 0.25 (Hertel 1966). Cruising speeds of 3-m-long bluefin tunas measured in large enclosures reached 1.2 L/s (260km/d), where L equals body length (Wardle et al. 1989). Angelfish (Pterophyllum scalare) and butterfly fish (Chaetodon sp.) (Fig. 1.3d) are maneuvering experts with short bodies and high dorsal and anal fins. Species of this guild live in spatially complex environments. Coral reefs and freshwater s...