
- Covers a variety of topics fundamental for music perception, including musical syntax, musical semantics, music and action, music and emotion
- Includes general introductory chapters to engage a broad readership, as well as a wealth of detailed research material for experts
- Offers the most empirical (and most systematic) work on the topics of neural correlates of musical syntax and musical semantics
- Integrates research from different domains (such as music, language, action and emotion both theoretically and empirically, to create a comprehensive theory of music psychology
eBook - ePub
Brain and Music
About this book
A comprehensive survey of the latest neuroscientific research into the effects of music on the brain
Trusted by 375,005 students
Access to over 1 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Part I
Introductory Chapters
1
Ear and Hearing
1.1 The ear
The human ear has striking abilities of detecting and differentiating sounds. It is sensitive to a wide range of frequencies as well as intensities and has an extremely high temporal resolution (for detailed descriptions see, e.g., Geisler, 1998; Moore, 2008; Pickles, 2008; Plack, 2005; Cook, 2001). The ear consists of three parts: The outer, the middle, and the inner ear. The outer ear acts as a receiver and filters sound waves on their way to the ear drum (tympanic membrane) via the ear canal (meatus), amplifying some sounds and attenuating others (depending on the frequency and direction of these sounds). Sound waves (i.e., alternating compression and rarefaction of air) cause the tympanic membrane to vibrate, and these vibrations are subsequently amplified by the middle ear. The middle ear is composed of three linked bones: The malleus, incus, and stapes. These tiny bones help transmit the vibrations on to the oval window of the cochlea, a small membrane-covered opening in the bony wall of the inner ear and the interface between the air-filled middle-ear and the fluid-filled inner ear (Figure 1.1).
Figure 1.1 Top: The major parts of the human ear. In the Figure, the cochlea has been uncoiled for illustration purposes. Bottom: Anatomy of the cochlea (both figures from Kandel et al., 2000).
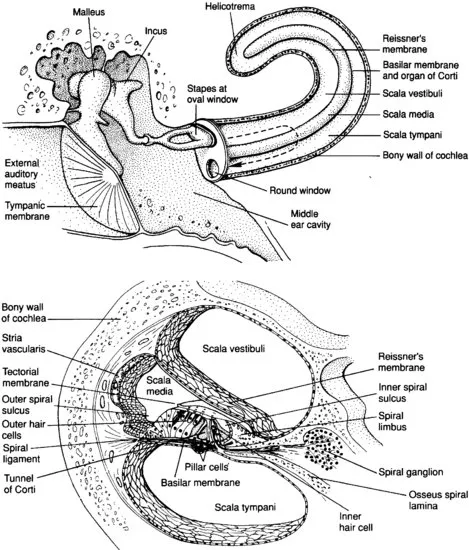
The cochlea has three fluid-filled compartments, the scala tympani, the scala media, and the scala vestibuli (which is continuous with the scala tympani at the helicotrema). Scala media and scala tympani are separated by the basilar membrane (BM). The organ of Corti rests on the BM and contains the auditory sensory receptors that are responsible for transducing the sound stimulus into electrical signals. The vibration of the stapes results in varying pressures on the fluid in the scala vestibuli, causing oscillating movements of scala vestibuli, scala media (including BM) and scala tympani (for detailed descriptions see, e.g., Geisler, 1998; Pickles, 2008).
The organ of Corti contains the sensory receptor cells of the inner ear, the hair cells (bottom of Figure 1.1). There are two types of hair cells, inner hair cells and outer hair cells. On the apical surface of each hair cell is a bundle of around 100 stereocilia (mechanosensing organelles which respond to fluid motion or fluid pressure changes). Above the hair cells is the tectorial membrane that attaches to the longest stereocilia of the outer hair cells. The sound-induced movement of the scalae fluid (see above) causes a relative shearing between the tectorial membrane and BM, resulting in a deflection of the stereocilia of both inner and outer hair cells. The deflection of the stereocilia is the adequate stimulus of a hair cell, which then depolarizes (or hyperpolarizes, due to the direction of deflection) by opening an inward current (for detailed information see Steel & Kros, 2001).
The inner hair cells then release glutamate (Nouvian et al., 2006)1 at their basal ends where the hair cells are connected to the peripheral branches of axons of neurons whose bodies lie in the spiral ganglion. The central axons of these neurons constitute the auditory nerve. The release of glutamate by the hair cells excites the sensory neurons and this in turn initiates action potentials in the cell's central axon in the auditory nerve. Oscillatory changes in the potential of a hair cell thus result in oscillatory release of transmitter and oscillatory firing in the auditory nerve (for details see, e.g., Pickles, 2008; Geisler, 1998). The duration of an acoustic stimulus is encoded by the duration of activation of an auditory nerve fibre.
Different frequencies of sounds are selectively responded to in different regions of the cochlea. Each sound initiates a travelling wave along the length of the cochlea. The mechanical properties of the basilar membrane vary along the length of the cochlea; the BM is stiff and thin at the basal end (and vibrates more to high frequency sounds, similar to the high e-string on a guitar, which resonates at a sound frequency of ∼330 Hz), whereas at the apex the BM is thicker and less stiff (and resonates at sounds with lower frequencies, similar to the low e-string on a guitar, which resonates at a sound frequency of ∼82 Hz). Different frequencies of sound produce different travelling waves with peak amplitudes at different points along the BM. Higher frequencies result in peak amplitudes of oscillations of the BM that are located nearer to the base of the cochlea, lower frequencies result in oscillatory peaks near the apex of the cochlea (for more details see, e.g., Pickles, 2008; Geisler, 1998).
The outer hair cells specifically sharpen the peak of the travelling wave at the frequency-characteristic place on the BM (e.g., Fettiplace & Hackney, 2006). Interestingly, outer hair cells achieve the changes in tuning of the local region in the organ of Corti by increasing or decreasing the length of their cell bodies (thereby affecting the mechanical properties of the organ of Corti; Fettiplace & Hackney, 2006). This change in length is an example of the active processes occurring within the organ of Corti while processing sensory information. Moreover, the outer hair cells are innervated by efferent nerve fibres from the central nervous system, and it appears that the changes in length are at least partly influenced by top-down processes (such processes may even originate from neocortical areas of the brain). Therefore, the dynamics of the cochlea (determining the processing of acoustic information) appears to be strongly influenced by the brain. The dynamic activity of the outer hair cells is a necessary condition for a high frequency-selectivity (which, in turn, is a prerequisite for both music and speech perception).
Corresponding to the tuning of the BM, the frequency-characteristic excitation of inner hair cells gives rise to action potentials in different auditory nerve fibres. Therefore, an auditory nerve fibre is most sensitive to a particular frequency of sound, its so-called characteristic frequency. Nevertheless, an individual auditory nerve fibre (which is innervated by several inner hair cells) still responds to a range of frequencies, because a substantial portion of the BM moves in response to a single frequency. The sound pressure level (SPL, for explanation and medical relevance see, e.g., Moore, 2008) is then encoded (1) by the action potential rate of afferent nerve fibres, and (2) by the number of neighbouring afferent nerve fibres that release action potentials (because the number of neurons that release action potentials increases as the intensity of an auditory stimulus increases). The brain decodes the spatio-temporal pattern consisting of the individual firing rates of all activated auditory nerve fibres (each with its characteristic frequency) into information about intensity and frequency of a stimulus (decoding of frequency information is dealt with in more detail further below).
1.2 Auditory brainstem and thalamus
The cochlear nerve enters the central nervous system in the brain stem (cranial nerve VIII).2 Within the brain stem, information originating from the hair cells is propagated via both contra- and ipsilateral connections between the nuclei of the central auditory pathway (for a detailed description see Nieuwenhuys et al., 2008). For example, some of the secondary auditory fibres that originate from the ventral cochlear nucleus project to the ipsilateral superior olivary nucleus and to the medial superior olivary nucleus of both sides (both superior olivary nuclei project to the inferior colliculus). Other secondary auditory fibres project to the contralateral nucleus of the trapezoid body (that sends fibres to the ipsilateral superior olivary nucleus; see Figure 1.2). The pattern of contra- and ipsilateral connections is important for the interpretation of interaural differences in phase and intensity for the localization of sounds.
Figure 1.2 Dorsal view of nerve, nuclei, and tracts of the auditory system (from Nieuwenhuys et al., 2008).
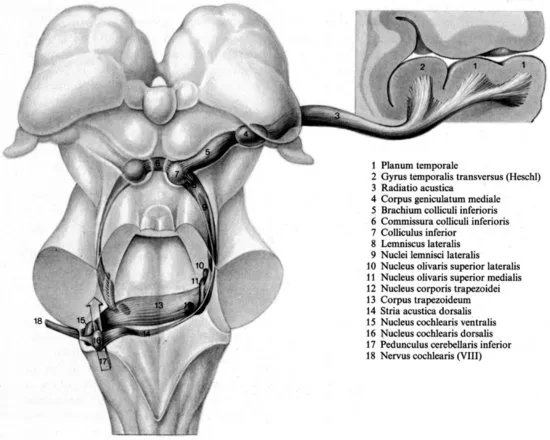
The inferior colliculus (IC) is connected with the medial geniculate body of the thalamus. The cells in the medial geniculate body send most of their axons via the radiatio acustica to the ipsilateral primary auditory cortex (for a detailed description see Nieuwenhuys et al., 2008). However, neurons in the medial division of the medial geniculate body (mMGB) also directly project to the lateral amygdala (LeDoux, 2000); specifically those mMGB neurons receive ascending inputs from the inferior colliculus and are likely to be, at least in part, acoustic relay neurons (LeDoux et al., 1990). The MGB, and presumably the IC as well, are involved in conditioned fear responses to acoustic stimuli. Moreover, already the IC plays a role in the expression of acoustic-motor as well as acoustic-limbic integration (Garcia-Cairasco, 2002), and chemical stimulation of the IC can evoke defence behaviour (Brandão et al., 1988). It is for these reasons that the IC and the MGB are not simply acoustic relay stations, but that these structures are involved in the detection of auditory signals of danger.
What is often neglected in descriptions of the auditory pathway is the important fact that auditory brainstem neurons also project to neurons of the reticular formation. For example, intracellular recording and tracing experiments have shown that giant reticulospinal neurons in the caudal pontine reticular formation (PnC) can be driven at short latencies by acoustic stimuli, most presumably due to multiple and direct input from the ventral (and dorsal) cochlear nucleus (perhaps even from interstitial neurons of the VIII nerve root) and nuclei in the superior olivary complex (e.g., lateral superior olive, ventral periolivary areas; Koch et al., 1992). These reticular neurons are involved in the generation of motor reflexes (by virtue of projections to spinal motoneurons), and it is conceivable that the projections from the auditory brainstem to neurons of the reticular formation...
Table of contents
- Cover
- Title Page
- Copyright
- Preface
- Part I: Introductory Chapters
- Part II: Towards a New Theory of Music Psychology
- References
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.4M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Brain and Music by Stefan Koelsch in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Neuroscience. We have over one million books available in our catalogue for you to explore.