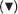Functional Biochemistry in Health and Disease provides a clear and straightforward account of the biochemistry that is necessary to understand the physiological functions of tissues or organs essential to the life of human beings. Focusing on the dynamic aspects of biochemistry and its application to the basic functions of the body, the book bridges the gap between biochemistry and medical practice.
Carefully structured within five sections, each biochemical, physiological or medical subject that is covered in the book is presented in one complete chapter. Consequently, each subject can be read and studied in isolation although cross-sectional links between the subjects are included where necessary. Background material, both biochemical and medical, that is necessary for an understanding of the subject, is included at the start of each chapter and clear, relevant diagrams enhance students' understanding. * Focuses on medically relevant aspects of biochemistry written from a physiological rather than a chemical perspective. * Clear presentation that minimises the use of jargon. * Each chapter contains boxes on related topics, relevant diagrams and a brief glossary. * Coverage includes athletic performance, apoptosis and the immune system. * Key historical developments are included to show how modern biochemistry has evolved.
By linking biochemistry, medical education and clinical practice this book will prove invaluable to students in medical and health sciences, biomedical science and human biology taking an introductory biochemistry course. In addition it will appeal to biochemistry and biology students interested in clinical applications of biochemistry.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Physical Activity: In Non-Athlet Athletes and Patients
In my opinion exercise physiology is from these viewpoints particularly important because an exercise situation in various environments provides a unique opportunity to study how different functions are regulated and integrated. In fact, most functions and structures are in one way or another affected by acute and chronic (i.e., in a training program) exercise. Therefore, exercise physiology is to a high degree an integrated science that has as its goal the identification of the mechanisms of overall bodily function and its regulation. It is regrettable that so few pages in standard textbooks of physiology for medical students are devoted to discussions of the effects of exercise on different functions and structures. That may explain why a majority of physicians do not recommend regular physical activity!
(Ästrand, 1997)
Football is the most popular sport in the world with approximately 40 million organised players (and c.100–150 million participants in total) … In football maximum potential needs to be activated in aerobic capacity, anaerobic alactacidpower, aerobic regenerative capacity … repair mechanisms, creativeness, intelligence and personality.
(Ekblom, 1994)
The term post-operative fatigue describes the state of overall drowsiness felt by patients who have undergone surgery: it can last for many days after the operation.
(McGuire et al., 2003)
Movement is an essential component of life. If something moves, it is assumed to be alive. External movement depends upon skeletal muscle whereas internal movement depends upon smooth and cardiac muscles. The latter muscles are discussed in other chapters. The term ‘physical activity’ is another term for external movement. Hence, it depends on skeletal muscle. This chapter contains a discussion of the biochemistry of skeletal muscle, including its involvement in everyday activities. This leads into the biochemistry underlying both the benefits and hazards of physical activity. In addition, the biochemical bases of the common problem of fatigue and the debilitating condition of chronic fatigue are discussed.
The mechanical basis of movement by skeletal muscle
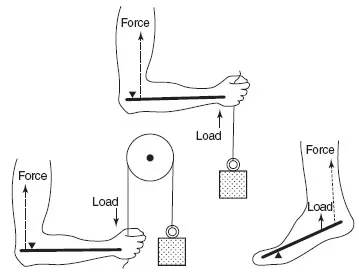
When a force acts on a rigid bar on one side of a fulcrum, a load can be moved on the other side of the fulcrum; this arrangement is known as a lever. Muscle and bone form a lever: the muscle generates the force, the fulcrum is the joint and the rigid bar is the bone (Figure 13.1). It is the process of shortening of a muscle, also termed contraction, that generates the force. However, for movement to occur, the process of contraction must be repeated in a cyclic fashion to provide for movement so that a muscle must lengthen as well as shorten. When a muscle is relaxed (i.e. not contracting) it can be extended (lengthened) by some force, usually due to the operation of an opposing muscle. In the upper arm, for example, the biceps extend the triceps and the triceps extend the biceps (Figure 13.2). Coordination between these muscles is essential to ensure smooth movement. Failure can result in one muscle tearing the other.
Contraction does not necessarily imply shortening but refers only to activation of the force-generating process. In order for shortening to occur, the force generated by a muscle must be greater than the force opposing the shortening. If the two forces are equal, there will be an increase in tension without shortening and the contraction will be isometric. It is also possible to increase the length despite development of tension, a situation known as ‘eccentric contraction’ (Table 13.1). The body contains approximately 600 skeletal muscles that comprise about 40% of normal body weight. Nonetheless it must be appreciated that muscle plays roles other than movement. These include the supply of amino acids by breakdown of muscle protein in some conditions to provide fuels for other tissues and synthetic processes (including glucose formation during starvation). Its ability to take up considerable amounts of glucose plays a significant role in the regulation of the blood glucose concentration after a meal. It also synthe-sises and stores glutamine, an important fuel for the immune and other cells.
Figure 13.1Muscle and bone as a lever. The figure shows the arm and leg as levers. The triangle
indicates the position of the fulcrum.
Figure 13.2Antagonistic muscles: their role in movement. The action of antagonist muscles is exemplified by the actions involved in delivering a punch. The arm is first /exed by contraction of the biceps muscle and simultaneous relaxation of the triceps muscle. The arm is then rapidly extended by contraction of the triceps muscle and relaxation of the biceps muscle. Such movements involve considerable nervous coordination.
Table 13.1 Different muscle actions
Eccentric contraction can most readily be appreciated by holding a weight in one hand close to the shoulder with the arm bent and lowering the arm from the elbow; the biceps are developing tension to hold the weight but increasing in length as the arm is lowered.
The versatility of muscle and the power output that can be achieved is demonstrated by athletic feats performed by many animals. The migratory bar-tailed godwit flies almost 6500 miles non-stop in 7 days, the monarch butterfly can fly 2000 miles at an average speed of 75 miles per day and mature eels make the epic journey from the Sargasso Sea to the rivers of Europe to spawn – and back again to die. Sprinters are also found in the animal world. A cheetah can accelerate to 45 miles per hour in 2 seconds and reach 70 miles per hour during the chase. Although all these athletic performances far outstrip those of humans, it must not be forgotten that humans are all-purpose animals. Individuals of this species have sprinted 100 m in under 10 seconds, run 5110 miles in less than 107 days (Max Telford ran from Anchorage, Alaska, to Halifax, Nova Scotia, in 1977), flown for 1 mile (with the aid of artificial wings but no additional power) and swum 21 miles nonstop across the English Channel. In addition, many human activities, such as playing a musical instrument, typing a manuscript or striking a moving ball, demand great precision of movement and an extraordinary coordination between muscles and the brain, which is achieved by no other single species.
Structure of muscle
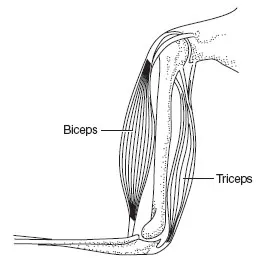
A muscle consists of groups of muscle bundles that join into a tendon at each end. The muscle bundles in the quadriceps are the vastus medialis, rectus femoris, vastus inter-medialis and vastus lateralis. Each bundle is separately wrapped in a sheath of connective tissue. Each muscle is composed of many fibres, packaged into bundles of about 1000 fibres (Figure 13.3). In some muscles (e.g. the psoas) the bundles run parallel to the long axis of the muscle. These are known as strap muscles and they contract quickly. In the muscles of the lower limbs and trunk, the bundles are arranged in a chevron fashion, oriented at an angle to the length of the muscle. These muscles are known as pennate. They develop greater tension but they contract more slowly than strap muscles.
Figure 13.3The arrangement of capillaries, fibres and bundles in a muscle.
In a strap muscle there is little connective tissue within the muscle since most is concentrated in tendons at the end. Such muscles produce the tenderest meat: fi llet steak is the psoas muscle of a cow. Pennate muscles contain more connective tissue within the muscle itself and hence form the cheaper cuts of meat.
Long, cylindrical fibres within the bundles correspond to the cells of the other tissues. These muscle fibres vary enormously in length from about 0.4 to 10–15 cm and in diameter from about 0.01 to 0.1 mm. Each fibre contains mitochondria and many nuclei (since it develops from a number of cells). It is surrounded by a plasma membrane (the sarcolemma).
A fibre is packed with longitudinally arranged myofi-brils that contract. This is the contractile unit of the fibre; it is 1–3 |jm thick in diameter. Each myofibril consists of about 1000–2000 filaments, which are known as myofilaments.
Muscles are attached to bone by tendons: tough, strong strands of connective tissue which transmit the force from muscle to bone. In addition, because of their elastic properties, they convert some kinetic energy into potential energy which is then used during the next stroke. This reduces the amount of ATP and hence the amount of fuel required to power running (Box 13.1).
Muscles differ widely in both their physiological properties (e.g. speed of contraction) and their metabolic properties (e.g. oxidative or glycolytic capacities). Originally, muscles were distinguished by the depth of their red colour, caused by the contents of myoglobin and cytochromes. Red muscles contain more myoglobin, which binds oxygen in the cell, and more cytochromes indicating that ATP is generated largely from oxidation. White muscle contains much less myoglobin and cytochromes and relies on glycolysis for ATP generation. Redness, however, is a poor indicator of the biochemical and physiological characteristics, although the terms red and white are still occasionally used. Since muscles contain fibres that differ physiologically and biochemically, it is more useful to classify fibres rather than whole muscles.
Fibre types
Fibres in human skeletal muscle are classified into two types: I and II. The Type I fibres are oxidative and generate force more slowly than type II fibres. Type II fibres have a lower oxidative capacity (relying more on glycolysis to generate ATP), a higher myosin ATPase activity and a greater power output. Type II fibres can be further subdivided into type IIA and type IIB. Type IIA are somewhat similar to type I and generate most of their ATP from oxidative processes but glycolytic capacity is still moderate. In contrast, type IIB fibres generate ATP almost exclusively from glycolysis and have the highest power output of any fibre although this can be maintained only for short periods ( i.e. explosive muscles) (Table 13.2). Fibre types can be identifi ed by histochemical methods performed on a biopsy sample (Appendix 13.1). Microtechniques allow individual fibres to be isolated and studied so that, for example, fuel contents and enzyme activities can be measured. The different fibres have different properties, and different proportions are present in different individuals. The biochemical and physiological characteristics of the different fibres explain why some individuals are good sprinters whereas others are good en durance athletes. These differences are most marked in top-class (elite) athletes. Top-class sprinters have over 70% of type II fibres, whereas top-class long distance runners have a high proportion of type I fibres (Table 13.3 and Figure 13.4). A similar trend is seen in domestic animals: horses bred for racing have a very high proportion of type II fibres; the heavy hunters, which are bred for their stamina, have a high percentage of type I fibres.
The maximum rate of glycogenolysis in type II fibres, as measured by difference in glycogen levels, is identical to the maximum activity of the enzyme phosphorylase measured in extracts of the same individual type II fibres in vitro. The maximum in vitro activity of this enzyme provides an excellent indication of the maximal capacity for glycogen breakdown even in individual fibres.
If your muscles have an average fibre composition, there is no chance of you winning an Olympic gold medal in the marathon or the 100 metre sprint. Although the percentage of type I and type II fibres is largely inherited, training can result in type IIc fibres, which are uncommitted fibres, being converted to type IIA or type IIB according to the type of training.
Box 13.1 The Achilles tendon: springs in the leg
Much of the energetic cost of running on the fl at is expended against the pull of gravity since, at each step, the centre of gravity of the body is raised. Not all energy is lost, however. The transfer of potential into kinetic energy is the basis of the children’s toy, the pogo-stick. Similarly, the elastic components of the leg allow some of the energy used to raise the body’s centre of gravity to be stored as potential energy and transferred back to kinetic energy for use in the next stride. The Achilles tendon is the main spring, stretching by about 1.5 cm (6% of its length), as the foot hits the ground. Some of the kinetic energy in the stride is transferred to potential energy in the tendon. More than 90% of this potential energy is also transferred back to kinetic energy in the next stride. The arch of the foot is another spring. As it fl attens, it transfers some 15% of the kinetic energy into potential energy which is transferred back to kinetic energy in the next stride.
Kangaroos put the pogo-stick principle to particularly effective use and elite athletes are much more ‘kangaroo-like’ than non-elite (see Figure 13.B). It has been calculated that is to these springs the effect of raise the effi ciency of conversion of the chemical energy into mechanical energy from about 25% to more than 40%, a remarkable increase. It is unclear iftendon elasticity can be improved by training. Unfortunately, with ageing, this energy-recycling property of the tendons is progressively lost.
It is this principle that explains why a rubber ball travels further by bouncing. It is well known, however, that a ball does not bounce well in a fi eld of mud. Nor do elite track athletes. Steve Ovett,* a world record holder for the mile and 1500 m races, illustrated this point when he described his performance in a cross-country championship early in his career (1970):
The race was a disaster. It was pouring with rain all day and by the time my event started the course was churned into a quagmire and at one point looked like a river. I am a fl uid runner and this was a course for sloggers … It was desperately tough … I think I fi nished twenty-seventh!
(Ovett & Rodda 1984)
Figure 13.B In the hopping kangaroo, the effect of the long Achilles tendon is that faster locomotion does not use more ATP! Indeed, the faster the kangaroo hops the more efficient it becomes in relation to use of chemical energy. In contrast, the kangaroos when walking follows the usual response to a faster pace, more ATP is used. As the kangaroo lands, the tendon is stretched, converting kinetic energy into potential energy. The cheetah is also eco...
Table of contents
Cover
Title
Copyright
Dedication
Preface
Acknowledgements
Abbreviations
I: INTRODUCTION
II: ESSENTIAL TOPICS IN DYNAMIC BIOCHEMISTRY
III: ESSENTIAL METABOLISM
IV: ESSENTIAL PROCESSES OF LIFE
V: SERIOUS DISEASES
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Functional Biochemistry in Health and Disease by Eric Newsholme,Anthony Leech,Anthony Leech in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biochemistry. We have over 1.5 million books available in our catalogue for you to explore.