Plant Polysaccharides, an exceptional new volume in Wiley-Blackwell's successful Annual Plant Reviews series, covers the polysaccharides and proteins that form the fundamental architecture of the plant cell wall, and the genes that encode the cellular machinery that synthesizes them.
The volume focuses on the evolution of the many families of genes whose products are required to make a particular kind of polysaccharide, bringing attention to the specific biochemical properties of the proteins to the level of kinds of sugar linkages they make.
Beautifully illustrated in full colour throughout, this exceptional new volume provides cutting edge up-to-date information on such important topics as cell wall biology, composition and biosynthesis, glycosyltransferases, hydroxyproline-rich glycoproteins, enzymatic modification of plant cell wall polysaccharides, glycan engineering in transgenic plants, and polysaccharide nanobiotechnology.
Drawing together some of the world's leading experts in these areas, the editor, Peter Ulvskov, has provided a landmark volume that is essential reading for plant and crop scientists, biochemists, molecular biologists and geneticists. All libraries in universities and research establishments
where plant sciences, agriculture, biological, biochemical and molecular sciences are studied and taught should have copies of this important volume.
CELL WALL POLYSACCHARIDE COMPOSITION AND COVALENT CROSSLINKING
Stephen C. Fry
The Edinburgh Cell Wall Group, Institute of Molecular Plant Sciences, The University of Edinburgh, The King’s Buildings, Edinburgh EH9 3JH, UK
Abstract:
Genetics now potentially lets us modify the production, crosslinking and degradation of cell wall polysaccharides. There remains, however, the need to test experimentally whether intended modifications of polysaccharide metabolism have successfully been effected in vivo. Simple methods for this are described, including in-vivo radiolabelling, enzymic dissection (e.g. with Driselase) and chromatographic/electrophoretic fractionation of dissection products.
After an overview of polysaccharide chemistry, I discuss the structures and taxonomic distribution of wall polysaccharides in charophytes and land plants. Primary and secondary walls are compared.
The major wall polysaccharides are cellulose [microfibrillar β-(1→4)-D-glucan], pectins (α-D-galacturonate-rich) and hemicelluloses (lacking galacturonate; hydrogen-bonding to cellulose; extractable by 6 M NaOH at 37 °C). Land-plant pectins are anionic polymers built of about four glycosidically interconnected domains (homogalacturonan, rhamnogalacturonans I and II, xylogalacturonan). Hemicelluloses occurring in most/all land plants are α-xylo-β-glucans, β-xylans (including α-arabino-β-xylans, α-glucurono-β-xylans, etc.) and β-mannans (including α-galacto-β-mannans, β-gluco-β-mannans, etc.). Another hemicellulose [mixed-linkage β-(1→3)(1→4)-D-glucan) is confined to Equisetum and some Poales.
Other taxonomically restricted features of angiosperm primary walls occur in Poales (xylose-poor xyloglucans; feruloylated arabinoxylans); Solanales and Lamiales (characteristic xyloglucans); Caryophyllales (feruloylated pectins); and Alismatales (apiogalacturonan). I also summarize characteristic wall features of charophytes, bryophytes, lycopodiophytes, fern-allies and gymnosperms.
The making or breaking of a ‘crosslink’ (defined as an individual chemical bond, not a whole ‘tethering’ chain) may cause wall tightening/loosening. Covalent crosslinks include phenolic coupling products, uronoyl esters and amides, and borate diesters.
This chapter discusses the main cell wall polysaccharides of streptophytes – i.e. land plants (embryophytes, from liverworts to angiosperms) plus charophytes (a group of algae sharing many subcellular features with land plants). Primary structures (sequences of sugar residues) and covalent crosslinks are discussed. A brief introduction to the vocabulary of polysaccharide (‘glycan’) chemistry, and the sugar abbreviations used, is included in the legend to Fig. 1.1; these abbreviations are used throughout the text without further definition. Further details of secondary and tertiary structures of polysaccharides can be found in Chapter 14. A major theme in the present chapter is the application of simple analytical methods by which polysaccharides can be identified, characterized and quantified, and their metabolism monitored in vivo. Another theme is the taxonomic distribution of various wall polysaccharides. Works complementing this chapter include Brett & Waldron (1996), Fry (2000), Schols & Voragen (2002), Mort (2002), O’Neill & York (2003) and Obel et al. (2006).
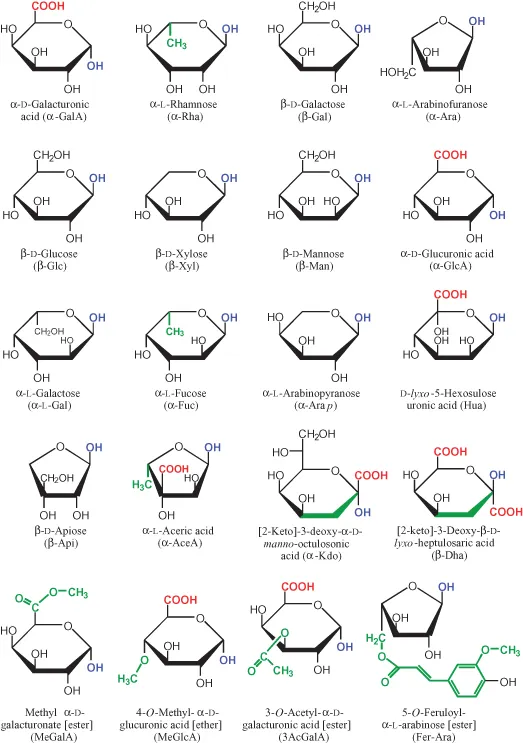
Figure 1.1 Monosaccharide building blocks (shown as Haworth formulae) of plant cell wall polysaccharides. The figure shows all the known sugar residues of plant cell wall polysaccharides and a selection of their esters and ethers. Top row, major components of pectins; 2nd row, major components of hemicelluloses; 3rd row, minor sugars of various origins; 4th row, mainly or only known from RG-II; bottom row, a selection of sugars with non-carbohydrate substituents. Sugars with five and six C atoms are called pentoses and hexoses respectively; Rha and Fuc are deoxyhexoses. The monosaccharides are shown as hemiacetal or hemiketal rings. However, within a polysaccharide, each sugar (except one, the reducing terminus) is present as an acetal or ketal residue, the term ‘residue’ implying that it is ‘what remains’ after losing the –OH group (shown in blue) from the anomeric carbon (the anomeric carbon is the one with single-bonds to two oxygens; it is here drawn as the right-hand extremity of the hexagon or pentagon). In a sugar residue of a polysaccharide, this particular –OH group has departed (in the form of H2O), ‘taking with it’ one oxygen-linked H atom from the next sugar along the polysaccharide chain. The one sugar of the polysaccharide that is not strictly a residue is the reducing terminus, so called because it has not lost its anomeric –OH group and in aqueous solution can therefore equilibrate with the straight-chain form, which possesses an oxo group (C=O, which has reducing properties). All but two of the sugars shown are aldoses (i.e. the anomeric carbon has only one additional C atom attached to it), but Kdo and Dha are ketoses (the anomeric C is attached to two other carbons). (Hua has two anomeric carbons (C-1 and C-5) and is both an aldose and a ketose.) In aqueous solution, each illustrated hemiacetal and hemiketal equilibrates with a small percentage of a straight-chain form possessing an oxo group (an aldehyde or ketone, in aldoses and ketoses respectively) – hence the slightly redundant term ‘keto’ in the names of Kdo and Dha. Each named sugar could theoretically occur as two isomeric forms (enantiomers, designated D- and L-), distinguished by the orientation of the C–O bond of the penultimate C atom. Galactose is the only wall residue known to occur as both D- and L-enantiomer. Note that D- and L-Gal differ in orientation of the C–O bond at all four non-anomeric, chiral centres (= carbons 2, 3, 4 and 5; the difference at C-5 is indicated by the placement of the –CH2OH group). The linkage between a sugar residue and the next building-block along a polysaccharide chain can be in either of two isomeric forms (anomers, designated α- and β-) defined by the orientation of the bond between the anomeric C atom and the oxygen atom (shown in blue) that bridges the two sugars: if this C–O bond has the same orientation as that of the penultimate C atom, then the residue is α-; if opposite, β-. This means that, in these Haworth formulae, the –OH of the anomeric carbon points down in α-D- and β-L-sugars, and up in β-D- and α-L-sugars. The sugar ring can be 6-membered (pyranose; -p) or 5-membered (furanose; -f). Api and AceA must be -f because of the absence of an oxygen on a C-5, and MeGlcA can only be -p. Ara occurs in both forms. All the others could theoretically occur in either form, but in practice occur only in the -p form illustrated. Each sugar residue is attached, via its anomeric carbon, to an –OH group on the following sugar unit in the polysaccharide chain. Usually, there are several such –OH groups to choose from (e.g., in the case of Glcp, on carbons 2, 3, 4 or 6: the linkage is designated (1→2), (1→3), (1→4) or (1→6), accordingly). However, a given sugar unit (either a residue or the reducing terminus) can and often does have more than one sugar residue attached to it. Once it has become part of a polysaccharide chain, a given sugar residue is ‘locked’ in one of the four possible ring forms (α-p, β-p, α-f, or β-f). These ring forms have a huge impact on the polysaccharide, as is obvious from the enormous differences in physical, chemical and biological properties between amylose and cellulose (which are α-p and β-p, respectively, but otherwise identical). Although illustrated here in unionized form, the free carboxy groups (–COOH, shown in red) would often be negatively charged (–COO−) under physiological conditions of pH. Relatively hydrophobic (non-polar) groups are shown in green. Abbreviations: The diagrams show (in parentheses) the shorthand used throughout this chapter. Thus, unless otherwise stated in the text, the ring-form (-p or -f) and enantiomer (D- or L-) are assumed to be as illustrated here; for example, ‘β-Gal’ implies β-D-Galp unless specified as L-Gal. Other abbreviations used (not illustrated): MeXyl, 2-O-methyl-α-D-Xylp (ether); MeFuc, 2-O-methyl-α-L-Fucp (ether); MeRha, 3-O-methyl-α-L-Rhap (ether); MeGal, 3-O-methyl-D-Galp (ether); 5AcAra, 5-O-acetyl-L-Araf (ester); 6AcGal, 6-O-acetyl-D-Galp (ester); 6AcGlc, 6-O-acetyl-D-Glcp (ester); ΔUA, a 4,5-unsaturated, 4-deoxy derivative of GalA or GlcA.
1.1.1 Some definitions
The best-known cell wall component is cellulose – a highly insoluble polysaccharide, of which the microfibrils (‘scaffolding’) of the wall are composed. Cellulose is, however, only one of many polysaccharides found in plant cell walls, usually accounting for less then half the wall’s dry mass. The other, non-cellulosic, wall polysaccharides (matrix components) are categorized into pectins and hemicelluloses.
1.1.1.1 Pectins
These were traditionally defined by their extractability from the wall with chelating agents, often with the assistance of heating (though this inevitably causes partial degradation of pectins and should not be used if a determination of molecular weight is planned) and often followed by ice-cold aqueous Na2CO3. A more acceptable definition of pectins is wall polysaccharides rich in α-GalA residues.
1.1.1.2 Hemicelluloses
These are not extracted by chelating agents or ice-cold Na2CO3, but are by concentrated aqueous alkali. They generally share the property of hydrogen-bonding to cellulose, at least in vitro; and some hemicelluloses probably also do this in muro, tethering adjacent microfibrils (Fry 1989; Hayashi 1989). For this reason, the term ‘crosslinking glycans’ was suggested for hemicelluloses. However, this term is not used here because the proposed in-muro tethering role remains largely hypothetical in many cases, and also because some non-hemicellulosic polysaccharides (e.g. rhamnogalacturonan II, RG-II) do crosslink.
1.1.1.3 Crosslinks
A ‘crosslink’, as the term is used here, is an individual chemical bond, e.g. an ester linkage or a hydrogen bond, that joins together two otherwise separate polymers; it is not a whole molecular chain that joins together two structures (e.g. a xyloglucan chain tethering two microfibrils).
Definitions of polysaccharide classes by their extractability from the wall are far from perfect. One problem is that some polysaccharides chemically identical to hemicelluloses but not hydrogen-bonded to cellulose can sometimes be solubilized with hot neutral water – for example, the bulk xyloglucan present in some seeds as ‘food reserves’. Another problem is that some hemicelluloses are covalently attached to pectins, resulting in hybrid polysaccharides that are difficult to classify. Nevertheless, the broad classification of wall polysaccharides into pectins, hemicelluloses and cellulose remains a useful convention.
1.1.1.4 Non-polysaccharide components
Also important in cell walls are non-polysaccharide components. First among these is water, typically accounting for around 60% of the wall’s total fresh weight, and around 70% of the fresh weight of the wall matrix (Monro et al. 1976). Water confers important physical properties on (hydrated) wall polysaccharides, acts as a solvent for apoplastic solutes and enables the functioning of wall-located enzymes. Changes in the water content of the matrix may expl...
Table of contents
Cover
Half title page
Title page
Copyright page
Series page
PREFACE
DEDICATION
CONTRIBUTORS
Chapter 1 CELL WALL POLYSACCHARIDE COMPOSITION AND COVALENT CROSSLINKING
Chapter 2 DISSECTION OF PLANT CELL WALLS BY HIGH-THROUGHPUT METHODS
Chapter 3 APPROACHES TO CHEMICAL SYNTHESIS OF PECTIC OLIGOSACCHARIDES
Chapter 4 ANNOTATING CARBOHYDRATE-ACTIVE ENZYMES IN PLANT GENOMES: PRESENT CHALLENGES
Chapter 5 BIOSYNTHESIS OF PLANT CELL WALL AND RELATED POLYSACCHARIDES BY ENZYMES OF THE GT2 AND GT48 FAMILIES
Chapter 6 GLYCOSYLTRANSFERASES OF THE GT8 FAMILY
Chapter 7 GENES AND ENZYMES OF THE GT31 FAMILY: TOWARDS UNRAVELLING THE FUNCTION(s) OF THE PLANT GLYCOSYLTRANSFERASE FAMILY MEMBERS
Chapter 8 GLYCOSYLTRANSFERASES OF THE GT34 AND GT37 FAMILIES
Chapter 9 GLYCOSYLTRANSFERASES OF THE GT43 FAMILY
Chapter 10 GLYCOSYLTRANSFERASES OF THE GT47 FAMILY
Chapter 11 THE PLANT GLYCOSYLTRANSFERASE FAMILY GT64: IN SEARCH OF A FUNCTION
Chapter 12 GLYCOSYLTRANSFERASES OF THE GT77 FAMILY
Chapter 13 HYDROXYPROLINE-RICH GLYCOPROTEINS: FORM AND FUNCTION
Chapter 14 PLANT CELL WALL BIOLOGY: POLYSACCHARIDES IN ARCHITECTURAL AND DEVELOPMENTAL CONTEXTS
Chapter 15 ENZYMATIC MODIFICATION OF PLANT CELL WALL POLYSACCHARIDES
Chapter 16 PRODUCTION OF HETEROLOGOUS STORAGE POLYSACCHARIDES IN POTATO PLANTS
Chapter 17 GLYCAN ENGINEERING IN TRANSGENIC PLANTS
Chapter 18 POLYSACCHARIDE NANOBIOTECHNOLOGY: A CASE STUDY OF DENTAL IMPLANT COATING
Index
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Annual Plant Reviews, Plant Polysaccharides by Peter Ulvskov in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.