![]()
1
Basic genetics
This chapter provides a review of basic genetics. It concentrates on the general principles that apply to normal, healthy animals. The exceptions to these principles are often the basis of genetic diseases, which are discussed in subsequent chapters.
Chromosomes
When a culture of rapidly dividing white blood cells is treated with the alkaloid colchicine (which halts cell division), and the cells are then stained and viewed under a light microscope, structures called chromosomes become clearly visible. They are scattered randomly within clusters, and each cluster contains all the chromosomes from just one cell. The area of genetics concerned with chromosomes is called cytogenetics.
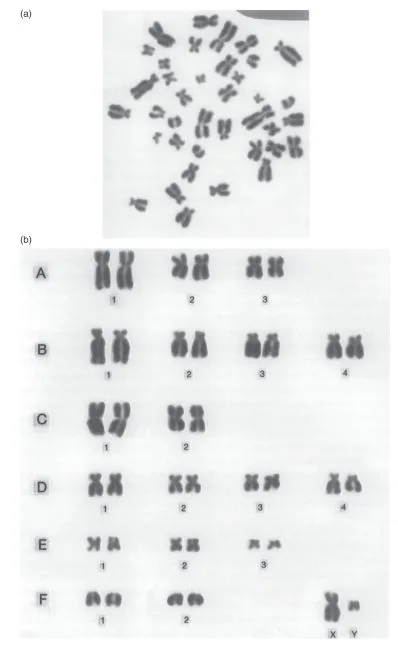
In order to study chromosomes more closely, a suitable cluster is chosen, as shown in Fig. 1.1a. Each item in the cluster consists of two rod-like structures joined together at a constricted point. Each rod-like structure is a chromatid and the constriction is a centromere. The two chromatids that are joined at the centromere have just been formed from one original chromosome. If the cell division had been allowed to proceed, the centromere would have split and each separate chromatid would then be called a new chromosome. For convenience, we talk of each pair of chromatids joined at the centromere as being just one chromosome, referring in fact to the chromosome that has just given rise to them.
All the chromosomes in the cluster are then rearranged in order of size. An arrangement such as this provides a picture of the complete set of chromosomes or karyotype of a cell (Fig. 1.1b). If many such arrangements are examined from normal, healthy individuals of both sexes of any species of mammal or bird, two facts become evident: each species has a characteristic karyotype and, within any species, each sex has a characteristic karyotype.
Karyotypes of different species differ in the shape, size, and number of their chromosomes. Within any species, all the chromosomes occur in pairs. In individuals of one sex, both members of each chromosome pair have the same size and shape. In the other sex, all but two chromosomes occur in such pairs, with the remaining pair consisting of two chromosomes of different size and shape. In this unequal pair, one chromosome has the same shape and size as members of one of the pairs in the opposite sex.
Fig. 1.1 (a) The chromosomes of a male cat, as seen through a light microscope. (b) The karyotype of a male cat, as obtained by rearranging individual chromosomes from (a). (Reproduced courtesy of P. Muir.)
The difference in karyotype between the two sexes is the key to sex determination. In mammals, the two chromosomes that form the unequal pair occur in males, and are called the X and Y chromosomes. In female mammals, one of the pairs of chromosomes consists of two X chromosomes. Thus in mammals, males are XY and females are XX. The X and the Y chromosomes are known as sex chromosomes. In birds, the sex chromosomes are given different names, and their relationship to sex is the opposite of that in mammals: male birds are ZZ and female birds are ZW. For convenience, we shall refer only to mammals in the following discussion, although all statements apply equally to birds if the names of the sexes are reversed.
Chromosomes other than the sex chromosomes are called autosomes. Within any species, males and females have the same set of autosomes, occurring in pairs. The sex chromosomes plus the autosomes constitute a genome, which is the total set of chromosomes in a cell. Genomes in which chromosomes occur in pairs are said to be diploid, and the two members of a pair are called homologues. In order to emphasize that chromosomes occur in pairs, the total number of chromosomes is called the 2n number, where n is the number of pairs. For example, the number of chromosomes in the karyotype illustrated in Fig. 1.1 is 2n = 38. To enable identification of each pair of chromosomes in a karyotype, the autosome pairs are labelled according to an internationally agreed convention, as shown in Fig. 1.1b. The two sex chromosomes are placed at the end.
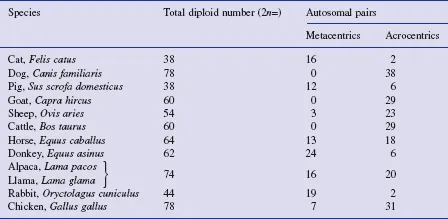
In order to describe karyotypes more fully, chromosomes are often classified according to whether the centromere is at one end (acrocentric), closer to one end than the other (sub-metacentric) or in the middle (metacentric). In this book, we shall follow common practice in using metacentric to cover both metacentric and sub-metacentric. The short arm of each chromosome is designated p (think of petite = small), and the long arm is designated q. (If the centromere is in the centre of the chromosome, the designation of which arm is called p is arbitrary, but is agreed by international convention; for acrocentric chromosomes, e.g. cattle autosomes, there is only one arm which, by convention, is designated the q arm). A summary description of the karyotypes of common domestic species is given in Table 1.1. Avian karyotypes are somewhat different to mammalian karyotypes, in that their smallest chromosomes are far smaller than the smallest mammalian chromosome. Until recently, chicken chromosomes were categorized as either macrochromosomes or microchromosomes, but now, in recognition of the continuum of sizes, they are classified into four groups, from group A (the largest chromosomes) to group D (the smallest).
Banding
When karyotypes were first investigated, individual pairs of chromosomes could be identified only according to their shape and size. Since then, various methods of staining chromosomes have been developed, giving rise to alternating light and dark regions called bands. The main types of bands are broadly classified as G (stained with Giemsa), Q (stained with quinacrine), R (reverse of G-banding), C (centromeres are stained), and T (telomeres are stained). Among specific banding strategies, we have G-banding by trypsin treatment and Giemsa staining (GTG), G-banding by early bromodeoxyuridine (BrdU) incorporation and Giemsa staining (GBG), R-banding by late BrdU incorporation and acridine-orange staining (RBA), and R-banding by late BrdU incorporation and Giemsa staining (RBG).
Table 1.1 A summary description of the karyotypes of some domestic species
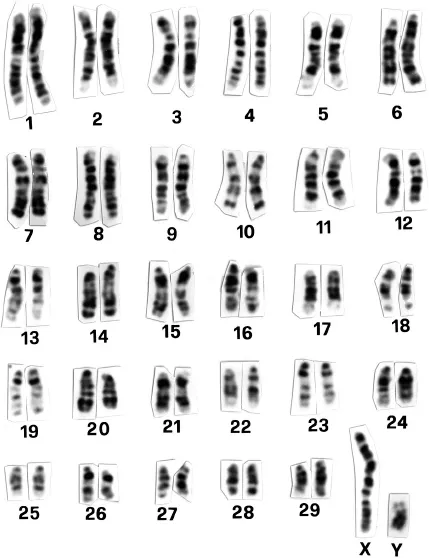
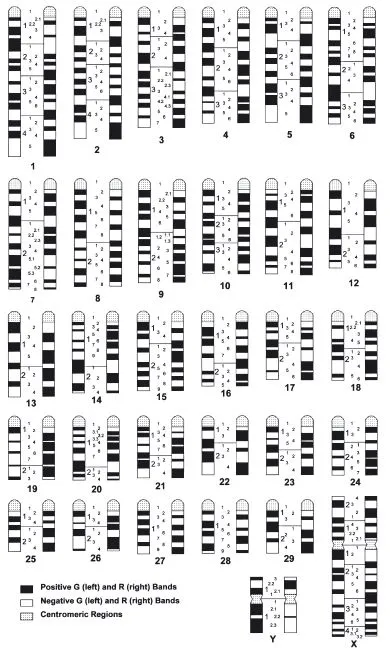
As an example of banding, the GBG bands of cattle are illustrated in Fig. 1.2. Since the position, width, and number of bands are different for each pair of chromosomes, each chromosome pair can be identified by its banding pattern. By studying many cells treated in the same way, it is possible to draw up an idiogram, which is a representation of the characteristic banding pattern for each pair of chromosomes. The bands are uniquely identified according to a convention known as the International System for Cytogenetic Nomenclature of Domestic Bovids (ISCNDB). Each arm is divided into a small number of regions which are numbered sequentially starting from the centromere. Then, in each region, the bands are numbered sequentially starting nearest the centromere. For example, the second band in the third region of chromosome 1 in cattle is designated q132, while the second band in the fourth region of the long arm of the X chromosome is Xq42. The ISCNDB idiogram for cattle is illustrated in Fig. 1.3. Banded karyotypes of other domestic species are illustrated in Appendix 1.1.
Meiosis and mitosis
For many thousands of years, humans have observed two phenomena in relation to sex determination in animals: first, that there is considerable variation in the numbers of each sex among the offspring of pairs of parents; and second, that despite this variation, the overall numbers of males and females across families are approximately equal.
Fig. 1.2 The standard GBG-banded cattle karyotype. (Reprinted by permission from S. Karger AG, Basel: ISCNDB (2000), Di Berardino, Di Meo, Gallagher, Hayes and Iannuzzi, Cytogenetics and Cell Genetics, 92, 283–99.)
As noted above, the difference in sex chromosomes between the two sexes is the key to sex determination. The reason why XX individuals are females and XY individuals are males is explained in Chapter 4. For the present, we shall ask simply: why is there so much variation in the numbers of XX and XY individuals in the offspring of pairs of parents, and yet at the same time approximately equal numbers of each sex overall? The answer lies in an understanding of gamete formation.
Fig. 1.3 The standard cattle idiogram, showing both G-bands (left) and R-bands (right). (Reprinted by permission from S. Karger AG, Basel: ISCNDB (2000), Di Berardino, Di Meo, Gallagher, Hayes and Iannuzzi, Cytogenetics and Cell Genetics, 92, 283–99.)
Meiosis
Meiosis is the process of gamete formation in which sperms are formed in testes of males and ova are formed in ovaries of females. The main result of meiosis is that each sperm and each ovum contains one member of each pair of chromosomes. Containing exactly one half of the usual diploid number of chromosomes, gametes are said to be haploid. The union of a sperm with an ovum at fertilization produces a zygote with the usual diploid number of chromosomes.
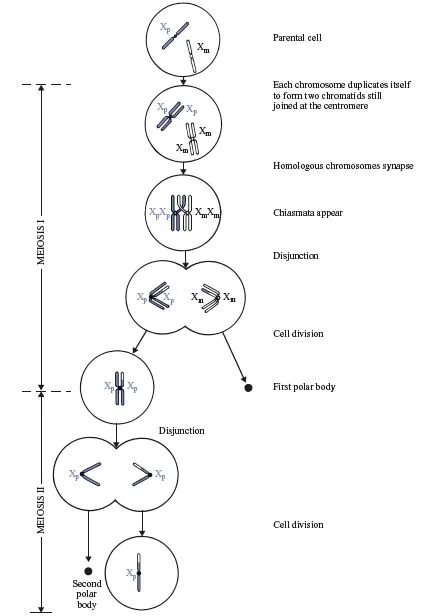
The process of meiosis commences with a normal cell containing the usual diploid set of chromosomes. To make the explanation easier, we shall consider what happens to just one pair of chromosomes (the sex chromosomes) in one sex (females), as illustrated in Fig. 1.4. In order to distinguish the two X chromosomes in females, we shall refer to them as Xp (paternal: originating from the father) and Xm (maternal: originating from the mother).
Meiosis occurs in two stages. Meiosis I begins with each chromosome duplicating itself, giving rise to two identical chromatids joined at the centromere. Then homologous chromosomes, in our case Xp and Xm, line up next to each other in the centre of the cell, in a process known as pairing or synapsis. This is facilitated by a protein structure called the synaptonemal complex, which ‘zips’ the two homologues together. The pair of homologues is called a bivalent. Because each chromosome has already duplicated itself into two chromatids, there are now four chromatids side by side in the cell; two Xp chromatids and two Xm chromatids. The two Xp chromatids are still joined at their centromere, as are the two Xm chromatids. At this stage, a process called recombination or crossing-over occurs, in which homologous chromatids each break at the same site and, in the process of re-uniting, exchange segments. This produces a cross-like structure called a chiasma (plural: chiasmata). In order to simplify the present discussion, we shall continue to refer to the chromatids as Xp or Xm, realizing that, as a result of crossing-over, any one chromatid may in fact consist of parts of both Xp and Xm. (A full discussion of the genetic implications of crossing-over is presented later in this chapter.) In the next stage of meiosis I, the two centromeres are pulled to opposite ends or poles of the cell, with the result that the two Xp chromatids move to one pole of the cell and the two Xm chromatids move to the other pole. Since this process involves the two pairs of chromatids disjoining from their previous paired arrangement, it is known as disjunction. In the final stage of meiosis I, the original cell divides into two cells; one contains the two Xp chromatids still joined at their centromere, and the other contains the two Xm chromatids, still joined at their centromere.
Following disjunction in females, only one cell continues to function normally; the other degenerates into a dark-staining structure known as the first polar body. It is entirely a matter of chance which of the cells remains functional. Consequently, there is an equal chance of either the two Xp chromatids or the two Xm chromatids ending up in the functional cell. (In Fig. 1.4, it happens to be the Xp chromatids that have survived.) In meiosis II in females, the two chromatids in the functional cell move apart (disjoin) and the cell divides into two cells, each containing one chromatid which is now called a chromosome. Once again, only one of the two cells remains functional; the other degenerates into the second polar body and, once again, it is entirely a matter of chance as to which of these two cells becomes the second polar body.
Fig. 1.4 Meiosis in a female, illustrated in terms of the sex chromosomes. A single cross-over has created two recombinant and two non-recombinant chromatids. The functional gamete in this example contains a recombinant chromosome with a paternal centromere. Exactly the same processes occur for all pairs of autosomes.
It is evident that in females, only one functional gamet...