The philosophy of Raymond Ruyer was an important if subterranean influence on twentieth-century French thought, and explicitly engaged with by figures such as Maurice Merleau-Ponty, Georges Canguilhem, Gilbert Simondon, and Gilles Deleuze. The Genesis of Living Forms is Ruyer's most focussed and forceful analysis of a central but apparently paradoxical biological phenomenon that also presents serious problems for philosophy: embryogenesis. When a cat develops from the early stages of fertilization to an adult, what is it that makes it the same cat? How is it that a living being can at once be the same and constantly changing?
Ruyer's answer to these questions unfolds through a detailed set of encounters with major scientific fields, from particle physics to social psychology, arguing that the paradox can only be dissolved by seeing the role that form plays in the ongoing development of living beings. In Ruyer's view, embryogenesis is a central problem not just in the life sciences; every thing must possess a relation to a form that is characteristic of it, from carbon atoms to embryos, and to embryologists themselves.

eBook - ePub
The Genesis of Living Forms
- 212 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Genesis of Living Forms
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Verticalism and Thematism
FUNCTIONING AND FORMATION
11 The fundamental feature of organic formations can be metaphorically characterised as a ‘verticalism’. Irreducible to function, formation can perhaps be said to be ‘perpendicular’ to it. A glance at the diagrams (figures 1.1–1.7) gets to the point more quickly than the explication that follows. Regardless of whether we consider the organic formation of a circulatory canal for air or blood, or an object of human creation, what is important is the opposition between the diagram’s vertical and horizontal arrangement. The first represents a formation, the appearance of new forms that nothing – except for the analogical knowledge [connaissance] of similar phenomenon – would allow us to deduce from the initial givens. The second represents the functioning of structures after their formation in space and time, a functioning which can easily be deduced by considering these structures. Every treatise on embryology includes a profusion of examples of vertical diagrams. Every treatise on physiology includes examples of horizontal diagrams.
12 In both cases, a given state orders the one that follows. The notion of the linking of forms is more general than that of functioning; there may be a linking together of forms without any functioning. There is never any pure emergence or appearance, in the sense in which we say that a ghost appears, or in which Venus appears above the stream ‘like a mist’. A gutter is formed from a flat panel, and the pipe is formed from the gutter.
PSEUDO-FORMATIONS
13 It is certainly the case that nothing but functioning takes place in a factory that manufactures gutters or pipes by stamping them out. On the other hand, when ocean sedimentation, viewed laterally, flows onto a continental shelf, forming folds which sometimes closely take on the appearance of a gutter – in short, what could be called the ‘morphogenesis’ of a mountain range – nobody would see anything but a mechanical function, and certainly nobody would have recourse to a mythology of a divine, ‘vertical’ and ‘artistic’ creation of mountains in anything but a religious or poetic sense. But organic morpho-genesis, even when it is very close to functioning, is something completely different because it results not only in the transformation of an initial form, and not only a brute increase in complexity that would be perfectly accounted for as a ‘quantity of information’, but in an increase in complexity in a self-sustaining, consistent, unified totality, capable of serving as the basis for a new formation in its turn. The brute increase in complexity in an open ensemble is not a sufficient criterion: it would probably take around the same number of words to telegraph a description of the Alps as it would to telegraph a description of mammalian embryogenesis. But once formed, the mountainous folds can only then function, that is, be passively subjected to pressure, erosion and chemical decomposition while the organism continues to be differentiated. Furthermore, at the moment of the formation of a mountain’s folds, we encounter the play of enormous exterior lateral forces on the sedimentary bed that press forward step by step. In the folding, invagination and migration of organic tissues, on the contrary, the forces in play must first be created or mobilised on the spot through a first differentiation. They are the ‘effects of explication’ as well as ‘explicative causes’, representing not
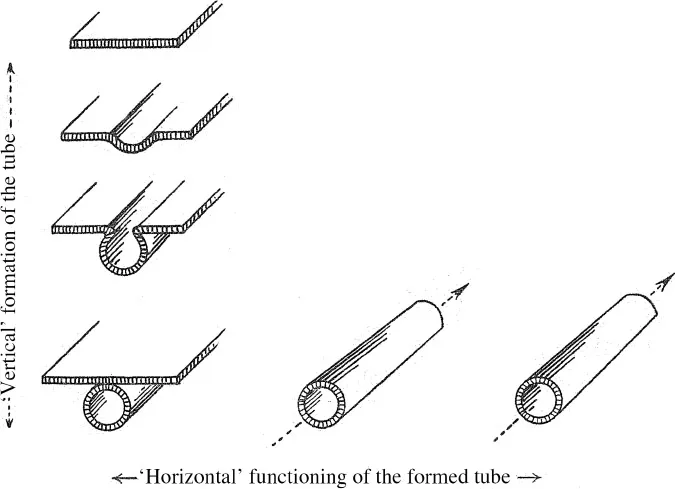
Fig. 1.1
an enormous dynamism but, to use Dalcq’s expression,1 an organisational dynamic – ‘organisational’ by virtue of the fact that it is already organised, distributed, hierarchised and synchronised.
There is an analogy between organic formation, as opposed to functioning, and the assembly of a new factory or production process involving new machinery, as opposed to the functioning of an established factory or processes of fabrication under way. Accelerated functioning increases production, but not the productivity of either the factory or the organism. Once established, the factory which fabricates gutters by stamping them out ‘functions’, but the assembly of a production process cannot be a functioning. It must have been an organisation, a true formation.
FORMATION OF THE AMPHIOXUS
14 Let’s recall here in broad strokes the early moments in the development of the amphioxus, which presents us with a kind of schema of development for all vertebrates. Without growing, the egg is segmented into two, then four, then eight cells of roughly equal size. Those cells that continue to divide constitute a small sphere the size of a blackberry (morula) before becoming a hollow sac (blastula).
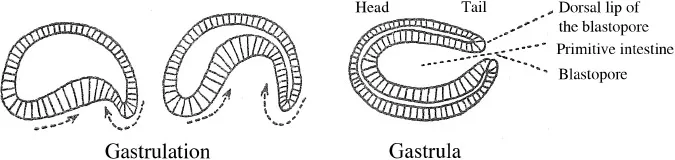
Fig. 1.2
15 The lower section flattens, collapses and subsides (gastrulation) into the higher hemisphere as if an invisible thumb were pressing on a rubber ball (figure 1.2). The cavity of the blastula is then reduced and a new cavity is formed, one which constitutes the primitive intestine. This primitive intestine communicates with the outside through the residual orifice: the blastopore. The area of the blastopore, above all on the dorsal side, is in fact the active agent of gastrulation. In the gastrula, the primordia of the animal have already been put in place. The gastrula has a cephalic-caudal axis with a bilateral symmetry and a dorsal-ventral axis. The parts which are invaginated, however, do not share the same destiny. The dorsal arch constitutes the chordal process,2 and in the mesoblast, the ventral side constitutes the endoblast which soon detaches itself from the mesoblastic primordia and is fused beneath the notochord (figure 1.3).
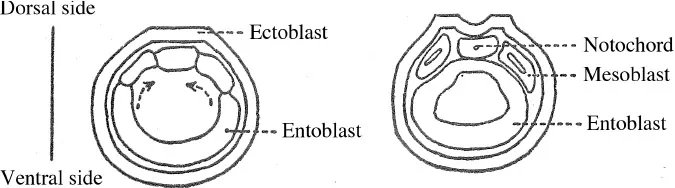
Fig. 1.3
On the dorsal side of the embryo, in contact with the notochord, the nervous system begins with two crests which appear along the length of the cephalic-caudal axis, bringing together and constituting, by fusing together, the neural tube. It must be noted that if, using the method of fate mapping,3 the cells which, once invaginated, will become the notochord, the mesoblast, or the intestine, can already be identified, this does not mean that they contain within themselves the structures in a preformed state for if these cells are experimentally displaced, they can yield something other than what they would have done had they been left in place and the embryo had developed normally.
The development of the amphibian egg is closely analogous but for the fact that being richer in reserved nutrient matter, segmentation and then gastrulation are somewhat impeded by this mass of vitellus and the dorsal side of the blastopore is rather more active (figure 1.4).
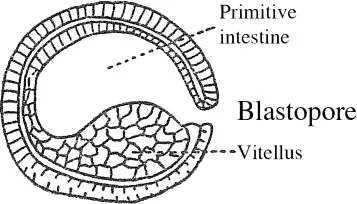
Fig. 1.4
Even in this rough-and-ready schematisation, it is difficult to overlook the characteristic nature of creative work, free behaviour, formation irreducible to functioning, or, in short, ‘verticalism’. In gastrulation, all is activity. If the blastula were a system in physical equilibrium, it is hard to see how its manner of pursuing the path that leads to such complication could be understood.
GRAFT AND PROSTHESIS
16 The progression and the transmission of forces or substances in horizontal functioning can be followed, allowing for the intervention by human technique and by way of more and more daring partial substitutions. We can already temporarily – and, soon no doubt, permanently – replace a failing organic canal with one made of plastic. In vertical formation, we can also discover chains of effects and the progression and transmission of substances in which intervention is possible. Experimental embryology presents us with techniques at least as daring as the techniques of prosthesis. In the precocious stages of a differentiation, for example, we can surgically transplant a graft from one part of the developing organism to another part, onto another organism of the same species or even onto an organism of a neighbouring species.
In a great many of these cases, normal development will nevertheless continue, the rootstock imposing on the graft its own mode of differentiation – here, for instance, contributing to the formation of a gutter (figure 1.5). Even in cases in which the graft does not completely conform to the inductions of the donor, developing as it would have in its original place, the two adjoined tissues at least partially harmonise with each other.

Fig. 1.5
17 But it is impossible to confuse these two kinds of technique, vertical and horizontal, graft and prosthesis. In a ‘vertical’ intervention, the surgeon counts on a competence possessed by the tissues involved. Through the displacement, he modifies the possibilities for the exercise of these competencies. He is the equivalent of a CEO who can modify the situation of his workers by allowing them to use their competencies to a greater or lesser extent; or, again, by modifying the signals that regulate their work, or modifying the rhythm according to which the materials they work on will arrive. He is not the equivalent of an engineer imagining machines that could substitute for the – already automatic – work of manual labourers or unqualified workers.
FUNCTIONING, MORPHOGENESIS AND BEHAVIOUR
Behaviour can be understood as a synthesis of functioning and formation. A machine, left to its own devices, functions. But when it is related to a theme or intended for a use, it has a ‘role’ that it ‘performs’. Behaviour, contrary to the thesis of mechanist behaviourists, is irreducible to a functioning. It implies improvisation and inventive adaptation to a function-role. In the hands of a pilot, an automobile, boat or plane behaves well or poorly depending on the pilot, whose intention thus takes the place of a vertical theme in a formation. A living organism, acting instinctively, also ‘behaves’. This is what signifies that a formative and improvisatory component which breaks with the functioning of the organs is already present: the ‘pilot’ is one with the machine. But at the same time, the pilot is an engineer whose improvised behaviour not only uses the machine but guides it, corrects it, completes and perfects it. Behaviour does not therefore depend exclusively on a structure – it is also the improvisation of structure. There is formation in all behaviour, and behaviour is the principle of formation.
18 In the higher animals, behaviour is related to the nervous system, which became the specialised organ of diffuse auto-conduction in the embryo. When a higher animal exhibits behaviour of some kind, its nervous system alone manifests the formative improvisatory component that is present in all behaviour while the rest of its body is constrained to function according to the guidance of the nervous system. In the behaviour of a protozoan, the entire organism at once functions and improvises. The unicellular organism improvises its pseudopods in order to move around, and its mouth and digestive tube in order to eat. In the embryo, too, even that of a higher animal, improvisation is everywhere implicated in the functioning of the organs already acquired through preceding developments. The blastula improvises – according to a specific theme, certainly, but beyond any automatic functioning – like the cell migration which transforms it into the gastrula, like the unicellular organism improvises its pseudopods. Behaviour is indiscernible from development, of which it is at once the principle and the manifestation. In the lower animals, bacterial colonies or amoebae, development remains indiscernible from behaviour until the death of the organism.
In an adult endowed not only with a nervous system but a brain with a developed cortex, as in a human being, it is extremely common for the whole of the organism to be represented multiple times by projection onto the cortex, like a sort of homunculus: sensibility is projected onto the parietal cortex; motricity, onto the frontal cortex; the visible body onto the occipital cortex; and even, in a still less precise fashion, sensibility and emotive motricity onto the pre-frontal cortex.4 In psycho-social behaviour, the functioning of the body is subordinated to the immediate behaviour of the ‘projected body’, of the cortical homunculus that behaves in the manner of a unicellular organism or an amoebic colony.
19 Consider a human being walking towards a determined goal on a poorly marked path requiring a certain inventive vigilance. The muscles of his limbs, insofar as they obey the impulsions of the nervous system, are equivalent to functioning. But the cerebral, sensori-motor homunculus, the zone from which nervous impulsions depart, cannot be limited to functioning: the cerebral homunculus, the ensemble of neurons woven into a network that constitutes it, forms a sort of enormous cerebral amoeba5 which ‘behaves’ according to an immediate auto-conduction analogous to the amoeba emitting pseudopods and completely transforming its apparatus as it moves. The voluntary locomotion of a human being is thus the functional amplification, obtained through complicated relays, of an improvised schema of movement in the homunculus of the motor cortex. In an adult multicellular organism, the brain is an apparatus which permits the separation of the ‘functioning’ component from the ‘behaviour’ component, which in the unicellular organism or the embryo merge together. The brain is restricted to functioning no more than the embryo is. This is why, for example, if a faradic current6 is applied to the motor zone of a living brain, variable motor ‘responses’ are produced rather than stereotypical movements. The cerebral homunculus is only local-isable precisely because it is behaving or in perpetual development, like an amoeba or a colony of amoebae in a crystalliser.7
Without succumbing to a vicious circle or a contradiction, we can assert that there is no difference between the claim that behaviour is a synthesis of formation and functioning (above all if we think of behaviour arising from a nervous system) and the claim that behaviour is the principle of functioning (above all if we think of beings that are not endowed, or not yet endowed, with a nervous system).
20 Should one wish to study the embryology of behaviour in the psychological sense of the word, as A. Gesell does, it becomes apparent that it is impossible to dissociate behaviour and morphogenesis.8 The thesis according to which biological heredity creates the organic structures which subsequently determine behaviour is today recognised as false. Heredity is at once a factor of structures and behaviours, or, to be more precise, formative behaviours and instinctive behaviours. More than a parallelism, there is an identity in nature between modes of development of the structures of the body and the modes of development of instinctive behaviours. ‘Vertical’ development is a behaviour, and instinctive behaviour develops in the fashion of an organic draft or primordium. The two are strictly interwoven, often through the convergence of fragmentary developments. In the ontogenesis of young passerines, the behaviour ‘scratching the head while balancing on a foot and a wing’ first appears in fragments. The parts of this complex movement appear before the behaviour, as a functional unity, can be accomplished. Ten-day-old marsupial foetuses (the opossum in particular has been studied by Hartman and McCrady), which resemble earthworms more than mammals, are capable of crawling from the urogenital canal to the pocket by using their anterior membranes and moving their heads from side to side until they arrive at the teat, where they attach themselves as if to an umbilical cord in order to complete their development. This behaviour makes use of several functions but envelops and inserts them in the developmental process as one phase amongst others. The sa...
Table of contents
- Note on the Translation
- Acknowledgements
- Introduction
- 1 Verticalism and Thematism
- 2 From the Molecule to the Organism
- 3 Internal Reproduction
- 4 The Fragmentation and Socialisation of Development
- 5 Signal Stimuli
- 6 Competence
- 7 Autonomous Procedures and Regulated Behaviour
- 8 Open Formations and Markovian Jargon
- 9 ‘Crossword’ Formations
- 10 ‘Spectacle-Spectator’ Complexes
- 11 Forms I, II and III
- 12 The Philosophy of Morphogenesis
- Notes
- Bibliography
- About the Author and Translators
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Genesis of Living Forms by Raymond Ruyer, Jon Roffe, Nicholas B. de Weydenthal, Jon Roffe,Nicholas B. de Weydenthal in PDF and/or ePUB format, as well as other popular books in Philosophy & Critical Theory. We have over 1.5 million books available in our catalogue for you to explore.