
- 168 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Chromosome Botany
About this book
The book covers both the basics of plant chromosomes as well as advances such as molecular characteristics, to give the reader an understanding of the chromosome as vehicle of hereditary transmission. It is meant for all levels of students in genetics, evolution, and biodiversity.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
CHAPTER 1
Chromosome: Structure and Components
Introduction
The term “chromosome”, coined by Baranetsky, is the principal component in the nucleus of higher plants, and is responsible for maintenance of hereditary stability. The chromosomes per se, containing the genes, form the material basis of heredity, the genes being arranged in a linear order. The chromosome complement of each cell contains the total gene content and constitutes the genome of the individual. The behavior of chromosomes follows a distinct pattern in the body cell or soma as well as in the germ cell line. The chromosome itself divides longitudinally into two halves, each half going to a daughter cell. Such equational separation of chromosomes as a whole, embodied in cell behavior termed mitosis, permits each and every cell of an organism to have same chromosome component and number. During the formation of gametes, responsible for sexual reproduction, the chromosomes adopt a different behavior. This behavior, otherwise known as meiosis, involves the coming together of the maternal and paternal chromosomes, which pair, forming bivalents, cross over and exchange segments followed by the two different chromosomes separating to two different poles. The daughter nuclei following such division contain half the number of chromosomes. Such nuclei with reduced number of chromosomes occurring on both the male and female side may go on multiplying to form gametes. During reproduction, the male and female gametes unite to form the zygote. After fertilization, the original number is restored. The continued equational division of zygote subsequent to embryo leads to the development of the adult individual, where all cells theoretically contain the same basic chromosome complement.
The process of mitosis and meiosis, i.e. equational and reductional division, ensures that every cell of an individual contains not only the same number of chromosomes but also all the genes of the complements.
Reductional separation leads to halving of the gene complements in the gamete. The fusion of the two gametes and subsequent development of fused cell restores not only the chromosome complements but also the total gene dosage. Exchange of segments occurs during crossing over – a phase of reduction division. In general, this type of behavior, universal for higher plants and animals, ensures uniformity and stability of the gene complement, in which chances of variability are inbuilt through the recombination mechanism and gene alteration.
In the basic chromosome structure, the chromonema thread is seen to have dense thickened areas with thinner regions in between, giving the appearance of a string of beads. The microscopic beaded regions are known as chromomeres and their position, and size as well, are constant to a certain extent.
Chromosomes at the cytological level have been regarded as composed of units of replication, transcription, mutation, and recombination.
Structural Details and Chemical Nature
In recent years, the structure and behavior of chromosomes have been analyzed in detail, aided by gradual advances in techniques. An understanding of the chromosome structure has been achieved at structural, ultrastructural, and molecular levels. The structural details are analyzed through different approaches, and these methods, combined with cytogenetic analysis, have helped in a better understanding of the basic structural pattern of chromosomes.
The eukaryotic chromosome at the molecular level represents a single long DNA molecule or several small DNA molecules forming a continuum, arranged end-to-end as several replicons or replicating units. Serial arrangement of DNA molecules also involves the assumption that linker sequences are present in between the different replicons or replicating units.
The chromosomes of higher organisms at the ultrastructural level have been shown to be made up of fibrils, approximately 20–30 Å in diameter, folded several times to yield a diameter of 100 Å thickness. It represents a continuous DNA–protein fiber in which condensed and decondensed segments alternate. A single condensed segment of a chromosome or chromomere of higher organisms is often comparable to the entire DNA skeleton of a microbe.
Uninemy
The earlier controversies with regard to strandedness of chromosomes have been fully resolved, and their uninemic nature has been confirmed.
The experiment of Taylor et al. (1957) using autoradiography had clearly indicated the uninemy of chromosome structure. This experiment involved treating dividing root tips of Vicia faba in nutrient solution with tritiated thymidine. Following replication, each daughter cell had each bipartite chromosome with one chromatid showing radioactive spots, indicating that each double helix in the chromatid had one parental strand and one new strand incorporating H3-thymidine. In subsequent replication, 50% of the chromosomes did not show radioactive spots. With subsequent divisions, lesser and lesser number of cells showed radioactive spots (Fig. 1.1). It has been claimed further that the DNA molecule is present along the entire length of the chromosome, of course associated with histones and other proteins. The molecule runs from telomere to telomere uninterrupted through the centromeric region. Although the amount of DNA per chromosome is very high, reaching the value of 1010–1011 daltons or even more in a single double-stranded DNA molecule, almost 2 m of this DNA can be confined within a nucleus less than 10 μm in diameter.
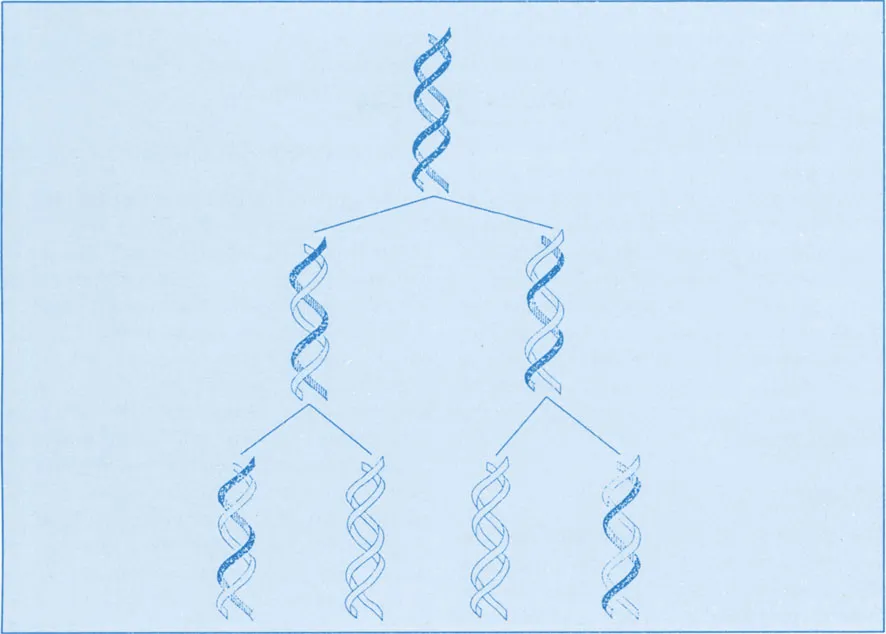
Figure 1.1 Schematic representation of the semiconservative mechanism of DNA replication.
Despite the presence of both histone and nonhistone proteins in chromosomes, DNA is responsible for maintaining structural integrity of the chromosomes. It is proved by the fact that treatment with DNAse alone can destroy the structural integrity of the chromosomes (Prescott, 1970).
Replicons
In this uninemic skeleton, chromosome structure represents a linear array of potentially independent replicons. The fact that the eukaryotic chromosome is made up of several multiple discrete units for replication has been specially demonstrated through DNA fiber autoradiography in cultured cells of mammalian tissue. The patterns of linear array of silver grains with decreasing density at both ends have been taken to suggest that replication is initiated at a central region of each unit and the chain elongation is bidirectional. As such, the chromosome structure is regarded as uninemic and multirepliconic in constitution. However, theories have been proposed regarding the presence or absence of linkers in replicons. Along the DNA fiber, the initiation of replication occurs at multiple internal sites. Taylor (1984) proposed a model presuming that the functional replication may be of the size of 100–300 kilobases, as indicated by autoradiography. A subset of the dispersed repeats may be responsible for suborigins in replicon at early development as well as during differentiation. Many of the suborigins may serve as sites for cessation of replication and initiation of transcription. The replication units represent a cluster of genes including some flanking regions.
Linkers
The presence of protein, RNA, and the divalent cations Ca++ and Mg++ indicates that these might serve as linkers. To test these hypotheses, enzyme digestion tests were performed and it was seen that neither protease nor nuclease digestion could disrupt the chromosome structure. Similarly, removal of cations through chelation, also left the structural integrity of the chromosomes undisturbed. These evidences show that the replicons in the DNA molecule are not attached to each other by means of any such linkers (Prescott, 1970). Linkers between the replicons may exist between two operons, i.e. through nonsense triplets or spacers.
The advantages of the replicon model are manifold. It provides a suitable base for independent replication of different chromosome segments. Thus, early and late replication can be easily accounted for. Similarly, the termination and initiation points of replicons may suggest loci for genetic crossing over. Translocation of segments, even reciprocal and at an intercalary level, can be explained through such a mechanism.
Heterochromatin and Euchromatin
Chromosomes are composed principally of two complexes – heterochromatin and euchromatin. The structural and functional differentiation of chromosomes into heterochromatin and euchromatin, spindle organizing regions, centromeres, chromosome ends or telomeres, and nucleolus organizing regions are well documented. Euchromatin constitutes the principal functional regions of chromosomes. Heterochromatin, on the other hand, is chiefly present on both sides of the centromere, at the telomeres, secondary constrictions, and also in a number of cases at the intercalary segments.
The term “heterochromatin” was originally derived from “heterochromosome” or sex chromosome, differentiating it from autosomes, by Heitz. Since sex chromosomes differ from rest of the chromosomes in their staining behavior, the chromosome segments showing a staining cycle similar to that of sex chromosomes were called heterochromatic. However, Darlington attributed to heterochromatin, the property of allocycly. It implies positive staining in chromosomes or heteropycnosis during interphase and reverse behavior in metaphase. However, in a number of cases, chromosomes may be apparently condensed throughout divisional cycle or decondensed throughout as in secondary constriction region. As such, all these segments were clubbed under the category “heterochromatin,” the common property being a staining behavior different from rest of the chromosome segments.
It was also presumed that genetically heterochromatin, though representing condensed state of chromatin, may be made up of genes having small, similar, and supplementary effects, as embodied in the concept of polygenes by Mather (1944). In later years, the discovery of repeated DNA sequences and their distribution in the chromosome indicated that heterochromatic segments are made up of repeated DNA, both GC/AT rich, the number and condensed state varying in different segments of chromosomes.
The term “heterochromatin” covers a rather heterogenous assemblage of chromatin represented by diverse characteristics in various species. In view of its repeat DNA constitution, nucleotypic effects have been attributed to heterochromatin. As regards function, variations on certain aspects of chromosomal metabolism, cytoplasmic synthesis, and nonspecific functions at the chromosomal level all categorized as “nucleotypic” are claimed to be regulated by these specialized segments, possibly influencing protein synthesis.
According to Vanderlyn (1949), any segment displaying properties different from those of euchromatin should be termed “heterochromatin.” This statement does not necessarily imply that all heterochromatic regions, including those of primary and secondary regions, are fundamentally alike. The general terms applied to heterochromatic regions earlier were constitutive or inherited and facultative or appearing during development.
Heterochromatic property may be manifested during development, the best example of facultative heterochromatin being provided by the mealy bug, where one set becomes heterochromatized during development and the other remains euchromatic. In sex chromosomes of mammals, due to dosage compensation, a single X remains active, the other becoming inactive or heterochromatized. Following cold treatment, certain regions on chromosomes of plants often show negative staining as in species of Fritillaria, Trillium, and Paris. Such areas, termed by Darlington as nucleic acid starved areas arising out of environmental changes, come also under the category of facultative heterochromatin.
Constitutive heterochromatin, on the other hand, remains stable in the chromosome and appears condensed during interphase as in prochromosomes in a large number of plant species such as Vicia faba. It is also often characterized by late or early replication of DNA, different from that of euchromatin, as in case of sex chromosomes. The best example of constitutive heterochromatin is provided by segments on both sides of the centromere, which remain as a single condensed block during metaphase, being designated as prochromosomes.
In addition to the centromere, constitutive heterochromatin is also located at the telomeric regions, nucleolar organizing regions, as well as intercalary position in certain organisms. The entire chromosome may also be heterochromatic, such as sex chromosome in some plants, as well as supernumerary or accessory chromosomes. At the entire chromosome level, B chromosomes or accessory chromosomes are mostly heterochromatic in nature. The number of such B chromosomes in a population may or may not vary. As compared to Allium stracheyii, where individuals differ in the number of B chromosomes, absolute constancy of such accessory chromosomes in all individuals of a population has been recorded in Tradescantia virginiana.
Variability of heterochromatin and its significance
Polymorphisms and genetic diversity
Constitutive heterochromatin shows variations both at the population as well as at the species level. Polymorphisms in chromosome structure at an intraspecific level have been recorded in several species of plants (Sharma, 1974). Such polymorphism may involve difference in nature and extent of constitutive heterochromatin in different individuals. This difference has been observed in characteristically stained bands appearing as Giemsa-stained regions. Such polymorphism in bands within homologous chromosomes has also been recorded, suggesting genetic diversity at the intraspecific level.
Within a genus, different species may show variable pattern of constitutive heterochromatin. Such variations have helped in tracing species relationship, evolution of karyotype, and role of structural alterations in evolution (Sharma, 1985).
Repeated DNA sequences, as mentioned earlier, are present in heterochromatin as large segments representing highly homogenous sequences. Heterochromatic regions in cereals often show differences in tandem array of repeated sequences, conferring variability of heterochromatin in different genotypes. Species-specific highly repeated sequences have been noted in heterochromatin of Alstroemeria aurea. The ribosomal RNA genes, viz. 18S, 26S, and 5S, are organized in high-copy tandem repeats and also show heterochromatic structure. Similarly, in pachytene chromosomes, rRNA genes have been located in tomato. In plant systems, variability of heterochromatin has been amply recorded as a criterion of genetic diversity.
In view of the presence of wide natural and induced variability in certain blocks of constitutive heterochromatin, it is presumed that a genome has a certain amount of tolerance so that parts of it can be lost or gained without any detectable phenotypic alterations. Also, more variation can be induced and be tolerated by cells. Therefore, studies on the variability in constitutive heterochromatin, coupled with its constitutive repetitive DNA in different cell populations of a species, may determine the tolerance of the genome for such variants and limits of this tolerance for more variability. These regions of constitutive heterochromatin may be interpersed within genes for essential sequences.
Role in amplification
Moreover, polymorphism in chromosome structure contributed by heterochromatic, segments shows quantitative DNA variations during organogenesis due to differential amplification of DNA. Such ontogenetic variations, arising out of differential amplification, have been recorded in several plants (vide Lavania, 1999; Nagl, 1978).
Researches have also revealed that chromosome constituents change their pattern during different phases of development, maintaining of course, a basic genetic make-up responsible for hereditary stability (Sharma, 1974, 1985). Chromosomes from such tissues where polytenic chromosomes are found (e.g. suspensor cells in plants or in tissues where the cell behaves endomitotically at the time of differentiation of organs), the distribution and amount of heterochromatin may be involved in the expression of character and organogenesis.
Role of recombination
In Nicotiana hybrids, exhibiting hybrid induced changes in chromosome morphology, evidences suggest that the ability to produce “megachromosomes” in N. tabacum (Gerstel and Burns, 1976) is the general property of heterochromatin expressed in an alien background.
Constitutive heterochromatin may help to recognize homologous pairing. Differences in heterochromatin content and C...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Table of Contents
- Preface
- Plant Chromosomes Today: An Introduction
- 1. Chromosome: Structure and Components
- 2. Chromosomes and Cell Division
- 3. Karyotype Concept
- 4. Chromosomal Changes and Biodiversity
- 5. Characteristics of Chromosome in Different Plant Groups
- 6. Chromosome Behavior in Differentiation
- 7. Nuclear DNA and Plant Evolution
- 8. Sex Chromosome Differentiation
- 9. Identification of Chromosome Segments
- 10. External Agents in the Induction of Gene and Chromosome Alterations
- Appendix
- Bibliography
- Subject Index
- Author Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Chromosome Botany by Archana Sharma in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.