
- 266 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Insect Reproduction
About this book
This book consisting of ten review chapters contributed by leading workers in their respective fields, from around the world, covers the whole subject of insect reproduction.It begins with the basic physiological questions of insect reproduction, moves on to discuss the new advances seen in the fields of behavioural and ecological mechanisms, and culminates by examining the recent work on evolutionary biology and its application in the field. Each chapter, although including a brief review of the basic seminal work, focuses mainly on the advances made within the last ten years and highlights those areas in which the respective authors see the greatest scope for further important advances
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Oogenesis and the Female Reproductive System
Contents
I. Introduction
II. Morphology of the Female Reproductive System
A. External Genitalia
B. Structure of the Ovary
C. Structure of the Female Accessory Reproductive System
1. Spermatheca and Spermathecal Accessory Glands
2. Colleterial Glands and Mesodermal Accessory Glands
3. Milk Glands
D. Endocrine Control of Differentiation of Accessory Glands and Ducts
III. Origin and Formation of the Germ Cells
IV. Oogenesis
A. Early Events in Oogenesis
1. Oocyte Differentiation
2. Endocrine Control of Early Oogenesis
3. Follicle Cell Differentiation
4. Trophic Function of Nurse Cells
B. Previtellogenesis
C. Vitellogenesis
1. Vitellogenin and Vitellin Chemistry
2. Vitellogenin Genes
3. Vitellogenin Synthesis
4. Vitellogenin Secretion
5. Uptake of Vitellogenin by the Ovary
D. Chorionization
1. The Vitelline Membrane
2. Chorion Formation
V. Ovulation and Oviposition
VI. Concluding Remarks
Reference
I. Introduction
Insect reproduction results from a succession of interdependent steps which are often quite different in nature and take place at various stages of the insect life cycle. The main reproductive events in females are sex determination, gonial mitoses and meioses, differentiation of the reproductive organs, previtellogenesis and vitellogenesis, accessory gland functioning, sexual behavior, mating, ovulation, and oviposition. It is the function of the female reproductive tract to produce the eggs and to deposit them at an appropriate time and in an appropriate place. In addition, the female reproductive tract must receive the spermatozoa from the male and transport them to the spermatheca where they are stored until they are used to fertilize the eggs as they are oviposited.
Insect reproduction strictly depends on environmental factors. Factors which may affect reproduction include temperature, humidity, photoperiod, nutritive conditions, and a suitable egg-laying substrate.
Regulation of insect reproduction involves numerous sensory receptors, nerve transmission, and integration in the brain, which regulates the synthesis of the two groups of insect developmental hormones, the juvenile hormones and the ecdysteroids, and produces its own neurohormones.1 The mechanisms which regulate each reproductive step may vary with insect species (see Chapter by Hardie).
Present knowledge of oogenesis has progressed variously, depending on the event considered. The best-known field is that of vitellogenesis and its endocrine regulation, while the early events of ovarian development and its “fine-tuning” control by hormones are less well understood.
Sexual reproduction is the general rule in insects, although many exceptions and modifications are observed (see Chapter by Dixon). In the more highly evolved social insects (Hymenoptera), reproduction is limited to a small number of individuals, often one queen and a small number of males (drones).
II. Morphology of the Female Reproductive System
The central site of egg production is the ovary. Ovaries are usually located dorsolateral to the gut and each comprises a number of tubular ovarioles ensheathed by a network of connective tissue (Figure 1). Each ovariole is composed of somatic and germ cell tissues. The number of ovarioles per ovary varies from one (e.g., in some viviparous aphids and in dung beetles) to about 3000 (in some higher termite queens).2 Three basic types of ovary organization are found in insects (Figure 2). The panoistic ovarian, probably the most primitive one, is found in the oldest families of insects, such as in Archaeognatha, Zygentoma, Odonata, Plecoptera, and in most orthopteroid insects, but also in some Megaloptera (Corydalidae) and most Siphonaptera. In the panoistic ovary, specialized nutritive cells are absent and most of the informational resources of the oocyte are provided by the synthetic activity of the oocyte nucleus itself. The polytrophic meroistic ovary is found in most endopterygotes, and in Dermaptera, Psocoptera, and in Phthiraptera. In polytrophic ovaries, a number of nurse cells are enclosed in each follicle along with an oocyte. In the Hemiptera, polyphage Coleoptera, Megaloptera (Sialidae)/Raphidioptera, and also in the most “primitive” winged insects, the Ephemeroptera,3 telotrophic meroistic ovaries are found. In this ovarian type, a syncytium of nutritive cells is connected with each oocyte by means of a trophic cord. In the vast majority of insect species, the nutrive, or yolk, contribution is supplied largely by the fat body, but in some cases the follicle cells can also serve as additional source of yolk. Accumulation of yolk (vitellogenesis; Section IV.C) normally occurs only in the terminal oocyte, that is, the oocyte closest to the oviduct. Another follicle cell function is the formation of the protective layers of the egg. These include the vitelline membrane and the chorion (Section IV.D).
Besides the ovary, oviduct-associated secretory cells can have important contributions to egg production (Section II.C). The most significant of such structures are the spermathecal accessory glands and the female accessory glands, such as the colleterial glands. Last, secretory functions of the vagina may play a role in egg production. Like other terrestrial animals, insects have had to solve the problem of bringing together sperm and egg in the absence of surrounding water (internal fertilization, see Chapter by Wall). Almost all insects store the spermatozoa received from the male in a specialized organ, the spermatheca, until they are used to fertilize the mature eggs.
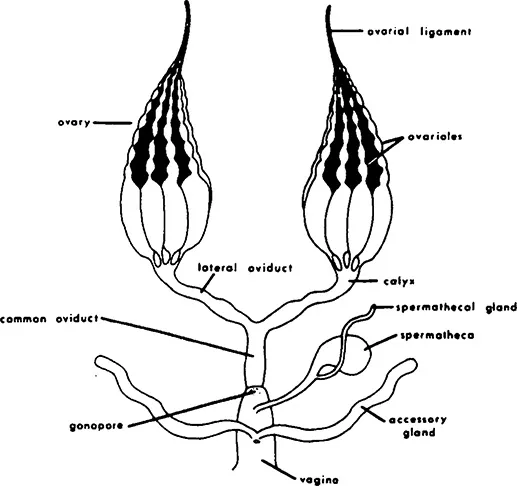
FIGURE 1. Female reproductive system: diagram of common type found in many insects. (From Gillott, C., Entomology, Plenum, New York, 1980, chap. 19. With permission.)
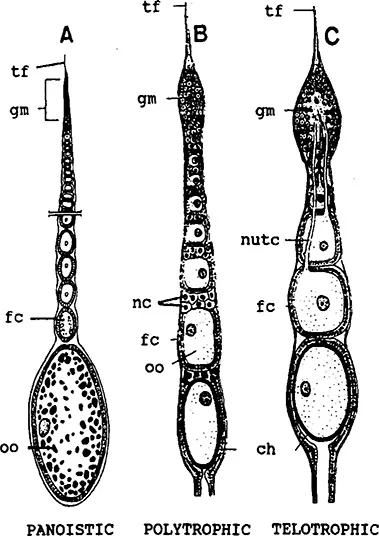
FIGURE 2. Schematic diagrams showing the types of ovarioles. Meroistic ovaries may be organized in the telotrophic and the polytrophic way, respectively.4,70 ch = chorion; fc = follicle cell; gm = germarium; nc = nurse cells; nutc = nutritive cord; oo = oocyte; tf = terminal filament. (From Gillott,C., Entomology, Plenum, New York, 1980, chap. 19. With permission.)
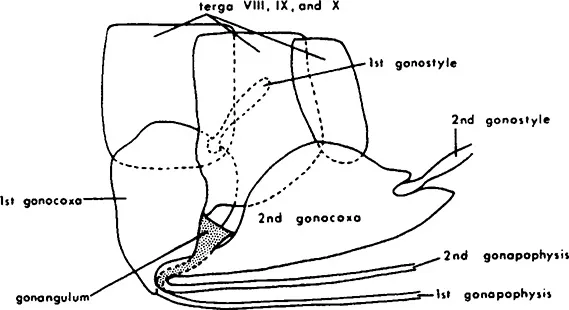
FIGURE 3. The primitive structure of the pterygote ovipositor in the thysanuran Lepisma. (From Gillott, C., Entomology, Plenum, New York, 1980, chap. 19. With permission.)
A. External Genitalia
The morphology of the organs specialized for copulation and oviposition is highly varied. In the mayflies, the oviducts open directly to two genital pores behind the seventh abdominal segment. In most insects, the appendages of the genital segments (eighth and ninth abdominal segment) form an ovipositor. In the apterygotes and some of the winged insects, the ovipositor is a simple opening both for copulation and for the deposition of eggs. The primitive structure of the pterygote ovipositor can be seen already in the thysanuran Lepisma (Figure 3).4 Among Pterygota, an ovipositor is found in Notoptera (Grylloblattodea), Dictyoptera, Ensifera, Caelifera, and Hymenoptera, some Odonata and most Hemiptera, Thysanoptera, and Psocoptera. The structure and elaborateness of the ovipositor is determined by the site of egg deposition. The ovipositor of Hymenoptera may be considerably modified for boring, piercing, sawing, and stinging. In the stinging Hymenoptera, such as bees, the eggs are released at the base of the ovipositor, and the ovipositor is modified by the addition of poison glands and reservoirs that evacuate the venom through the hollow sting. The ovipositor of Drosophila has sharpened ends that penetrate the surface of fruit, while the ovipositors of some of the predatory wasps are long (up to 15 cm in length) and sharp to penetrate the body of the insect prey.5
B. Structure of the Ovary
Insects have become particularly adept in manufacturing large numbers of oocytes within the ovary. The fruit fly, Drosophila melanogaster, can produce, during a 10-week reproduction period, a quantity of eggs equivalent to 30 times her body weight.6 Her newly formed oocytes can undergo a 100,000-fold increase in volume within 3 days. Such reproductive feats are possible because of certain evolutionary adaptations: (1) Oocytes have developed methods for incorporating massive quantities of female-specific proteins (vitellogenins) which are transported in the hemolymph from the fat body to the ovary. (2) Mechanisms have evolved for loading unfertilized eggs with the ribosomes, tRNA, and long-lived mRNA that are required in early embryogenesis. (3) The ovarioles are supplied by tracheae of the aeriferous type, with a great diversity in the methods by which oxygen is delivered to the individual oocytes.7
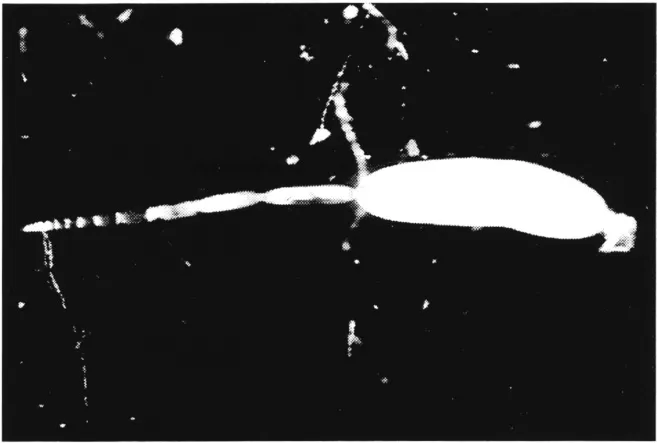
FIGURE 4. Panoistic ovariole of a cricket, Gryllus bimaculatus de Geer, with a mature egg in the terminal oocyte. Photograph courtesy of K. H. Hoffmann, Bayreuth.
Panoistic ovarioles can be developed by blocking germ cell cluster divisions totally, as is found in most “primitive” insects, or after germ cell cluster formation by final cleavage of cystocytes, all of which develop as oocytes as found in stone flies or thrips.8 In the panoistic ovary, each of the ovarioles is composed of a terminal filament, the germarium, a series of oocytes at the previtellogenic phase of development, one or more oocytes in the process of vitellogenesis, and last, the mature egg (Figure 2A; Figure 4). The terminal filament is made up of a group of flattened cells, surrounded by a basement lamina and an ovarian sheath, both of the latter surrounding the entire ovariole. The oogonia are located in the most anterior region of the germarium, followed by a zone of oocytes in the early stages of meiosis. At the posterior end of the germarium, the oocytes are beginning to be surrounded by a monolayer of follicle cells. The size increase of oocytes at the previtellogenic stage is accomplished by an expansion of the cytoplasmic volume. In many cases, a multilayered pad of interfollicular tissue is located between successive oocytes.9 In many insects with panoistic ovaries, vitellogenesis commences in the penultimate oocyte only after ovulation at the terminal one (e.g., in Locusta migratoria and Schistocerca gregaria [Caelifera]), or in the case of ovoviviparous cockroaches, after the loss of the egg case. The inhibitory effect is mediated by the interfollicular cells which pass an inhibitory substance from the anterior to the more posterior oocyte. In contrast, secretion from such cells located proximal to the oocyte stimulates vitellogenesis. In other species (e.g., in Periplaneta americana [Blattodea] and Melanoplus sanguinipes [Caelifera]), two or more oocytes may be vitellogenic at the same time, although at different stages of the yolk deposition cycle. All the above-mentioned insects produce eggs in batches. In the stick insect Clitumnus extadentatus, the different ovarioles can mature asynchronously, and the female lays a few eggs per day for several weeks. The number of ovarioles in panoistic ovaries can range from 4–3000 (5 in Acrididae, 15–30 in Tettigonioidea, 150–170 in Gryllidae, about 3000 in Isoptera queens).
A terminal filament and a germarium are also found in polytrophic meroistic ovaries (Figure 2B). In this case, the anterior region of the germarium contains one or more stem-line oogonia and a number of daughter cells or cystoblasts. The cystoblasts divide to give a cluster of cells remaining connected by structures called “ring canals” or “intercellular bridges.” The innovation of the polytrophic ovary is the differentiation of only one oocyte, which generates from one, central cell of the cluster, whereas all other siblings are transformed into nurse cells. In many cases, clusters follow the 2n-rule (1 oocyte + 2n – 1 nurse cells...
Table of contents
- Cover
- title
- copy
- Introduction
- The Editors
- Contributors
- toc
- Dedication
- Chapter 1 Oogenesis and the Female Reproductive System
- Chapter 2 Insect Male Mating Systems
- Chapter 3 Sex Determination in Insects
- Chapter 4 Hormones and Reproduction
- Chapter 5 Fatal Attraction: The Disruption of Mating and Fertilization for Insect Control
- Chapter 6 Parthenogenesis in Insects with Particular Reference to the Ecological Aspects of Cyclical Parthenogenesis in Aphids
- Chapter 7 Factors Affecting Fecundity, Fertility, Oviposition, and Larviposition in Insects
- Chapter 8 Protandry and Mate Acquisition
- Chapter 9 Swarm-Based Mating Systems
- Chapter 10 Male Nuptial Gifts: Phenotypic Consequences and Evolutionary Implications
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Insect Reproduction by S. R. Leather in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.