![]()
1
INTRODUCTION
The human brain is a mass of interconnecting neurons, the number of which is enormous. Estimates range from 1 to 100 billion neurons, and communications among the neurons are made possible by synapses, the number of which is orders of magnitude larger still. Meanwhile, the computer systems which are used to simulate brain functions contain fewer than 1 million elements and do not show the rich diversity of interconnections which is found in living organisms. This difference in scale between the actual brain and vaguely similar digital computers is so large that many of the mysteries of function, development and ageing of the real brain will undoubtedly remain mysteries for decades to come. Yet, having stated the obvious, it is nonetheless true that a few fundamental characteristics of brain function have been discovered and we can be confident of at least some of the structural pieces and functional relationships which must underlie a realistic theory of brain function. Moreover, such knowledge allows for computer simulations of the activity of small groups of neurons – making possible a second ‘experimental’ approach, in addition to that involving living brains.
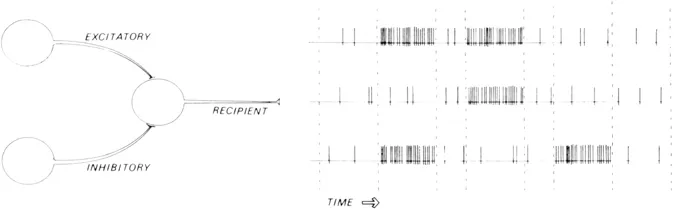
The first insight into brain function was undoubtedly the realization that indeed the brain is the biological organ which allows cognition. Our hearts and livers are important for other reasons, but the brain is the thinker. Secondly, the brain thinks with its massive network of neurons – not its glial cells, blood vessels or cerebrospinal fluid – and the thinking activity of the neurons is fundamentally electrical. The on-off electrical nature of neural activity has led to the familiar computer-brain analogy – the limitations of which we must keep in mind, but there is an important similarity between these two kinds of system: unlike analogies based upon clockwork mechanics or pumps and sluices, the computer analogy is not misleading insofar as neural activity is truly electronic. In detail, there are important differences, but in both cases we are talking about temporal and/or spatial patterns of electrical impulses which allow both systems to compute (see figure 1.1).
Two important facts about brain electronics should be noted. Each neuron is itself a unit which can receive various impulses, ‘digest’ that input and emit an output. How sophisticated the ‘decision-making capabilities’ of any one neuron may be remains uncertain, but each neuron is undoubtedly more complex than a small section of copper wire which merely relays all input. Each neuron summates the electrical information coming to it both spatially (over the surface of its dendrites, axons and cell body) and temporally (as a serial sequence of such inputs) and responds by exciting or inhibiting other neurons. Secondly, the neuron is unidirectional: the dendrites and axons which, generally speaking, act as the input and output channels to and from the neuronal cell body will normally allow pulses to travel in one direction only. Both of these established facts concerning neuronal function will prove to be important when we consider possible patterns of neural activity in the brain.
Figure 1.1 Neuronal inhibition and excitation. Here is shown a schematization of the effects of an inhibitory neuron and an excitatory neuron on a third neuron. Increased firing can lead to either a decrease or an increase in the firing of a neuron receiving the axonal output. As far as is now known, no other kinds of ‘Information’ are encoded in the temporal sequence of neuronal firing.
The brain code and the neuron code
An essential distinction between the neuron code and the brain code should be noted here. The neuron code is the proper subject matter of neurophysiology – and that code constitutes the fine-grained substrate of, but is not synonymous with, the brain code – which is the proper subject matter of neuropsychology in its broadest sense. On the one hand, the neuron code is concerned with the cellular mechanisms by means of which the electrical information in one neuron is summated with that of other neurons and then transmitted to still other neurons. On the other hand, the brain code is concerned with the relatively macroscopic mechanisms by means of which psychologically meaningful information (inevitably, multineuron patterns of activity) is transmitted from region to region within the brain.
Although brief discussion of the neuron code will be made in this and the next chapter, the primary concern of psychology and the topic of this book is not in fact the neuron code and the intricacies of cell biology. On the contrary, given certain known mechanisms of neuron-to-neuron communication, we will be interested in considering what kinds of mechanism may be involved as the brain juggles the thoughts and images which we know to be important in normal cognition. For the most part, we will let the cell physiologist worry about which neurotransmitters and how many synapses are involved, and we will concentrate on the relatively macroscopic dynamics of the brain. Again this is a topic for later chapters, but this recurring theme is worth stating clearly at the outset: the results and conclusions of biochemistry, molecular biology and neurophysiology will necessarily provide most of the bricks and mortar of a theory of the brain, but such building blocks are neither equivalent to nor do they add up simply and obviously to the neuronal cathedral which we are trying to understand. Provided with a firm grasp of the well-established aspects of neuron science, still our work in unraveling the puzzle of the brain has just begun.
Fortunately the bricks-and-mortar science of the brain has advanced remarkably over the past three decades. Most importantly, it has become possible to record the electrical activity of single cells in the living brain. Such single-cell recording experiments have shown unequivocally that one important aspect of the neuron code is simple firing rate. A given neuron in primary visual cortex, for example, will fire more frequently when a visual stimulus has certain (location, intensity, wave-length and/or orientation) characteristics, and less frequently when one or more of the relevant features of the stimulus are changed.
As yet little is known about the significance of the actual temporal sequence of the firing of a particular cell (or the significance of the interburst interval and other higher-order temporal features of the firing series). Especially as sensory information in various modalities is mixed together in association cortex, it may well be that more complex temporal codes related to the probabilities of simultaneous stimuli play an important role in abstracting high-level information from the sensory field. For example, the pattern of visual information which at some region of visual cortex is translated into the concept of ‘dog’ may be decoded as a neutral stimulus unless associated with the sounds of a dog barking. Depending upon the frequency with which this combination of stimuli has been associated with physical pain or other types of distress, these independently neutral stimuli may take on new meanings. In this example, it is the simultaneity of the auditory and visual stimuli which signals the potential danger.
In other words, the temporal sequence of neuronal firing is likely to be an important aspect of the neuron code in abstracting information from sensory stimuli, but it is noteworthy that the information abstracted from a temporal sequence is itself likely to be stored topographically (that is, spatially) on the cortex. The simplest manifestation of this process of translating the temporal dimension into spatial dimensions is the localization of high-frequency sounds anteriorly within primary auditory cortex, with lower frequencies located posteriorly. The actual firing rate of the cortical neurons involved does not affect the frequency of the perceived tone. Such ‘spatialization’ of the temporal dimension in the cortex is arguably one of the primary functions of the neuron code.
The detailed mechanisms involved in the neuron code will undoubtedly continue to fascinate the cell physiologist for decades to come, but the implications for the psychologist lie primarily in the fact that, however the brain code may work, it works in a world which is greatly simplified in terms of its temporal characteristics. The extremely high frequencies of photic stimulation, the lower frequencies of auditory stimulation and the extremely low frequencies of tactile stimulation are all translated into an artificial two-dimensional world where the on or off signals of neurons individually ‘mean’ stimulation of peripheral sense organs at a given frequency quite unrelated to the frequency of the cortical neuron firing.
The relationship between temporal and spatial encoding of sensory stimuli will be explored in later chapters. Suffice it to say that there are mechanisms for both temporal and spatial summation in the brain – some of which appear to be like the high-speed serial computations of computers and some of which appear to be like the topographical encoding of environmental stimuli on to a suitably sensitive ‘neuronal photographic film.’
Progress in determining the precise computational and sensory mapping mechanisms of the brain continues to be rapid. Currently several lacunae in our understanding of the cell physiology of the neuron code remain, but need not imply that elucidation of the fundamentals of the brain code will be impossible. Today we can be sure of the importance of at least two temporal aspects of the neuron code: the more frequent the firing of a given neuron, the more frequent or the less frequent the firing of those neurons receiving its axonal output. In other words, we can be sure of only excitatory and inhibitory relationships among neurons – the degree of the effect being a function of the firing rate. More complex things may in fact be happening between neurons, but we will see that already excitation/inhibition – unrelated to complex temporal factors – has direct implications for the brain code.
Evolution of the brain — the Vertical’ dimension
Climbing up the ‘tree’ of animal evolution, it is apparent that there are massive increases in the number of, and complexity of connections among, the neurons in animal brains. This is generally true in terms of the absolute numbers of neurons and is particularly striking in terms of the number of neurons relative to various measures of body size. But the essence of this increase in size and complexity is better understood in terms of the amount of nervous tissue interposed between nerve fibers carrying sensory input from the body surface and nerve fibers carrying motor output to effector organs. One such measure is the ratio of the size of the cerebral cortex relative to the number of ascending and descending nerve fibers – a better indicator than brain volume of the degree of brain activity intervening between stimulus and response. Thinking along these lines, various attempts at devising objective anatomical measures of the spectrum of animal intelligence have been made within the field of comparative neuroanatomy.
The underlying hope has been that if evolutionary changes in brain structure can be related to behavioral differences among species, then some progress in understanding how the brain thinks will be possible. It is surprising, however, how difficult it has proven to obtain a simple measure of the brain which reflects the phylogenetic development of a species – much less a measure which reflects intelligence or specific behaviors.
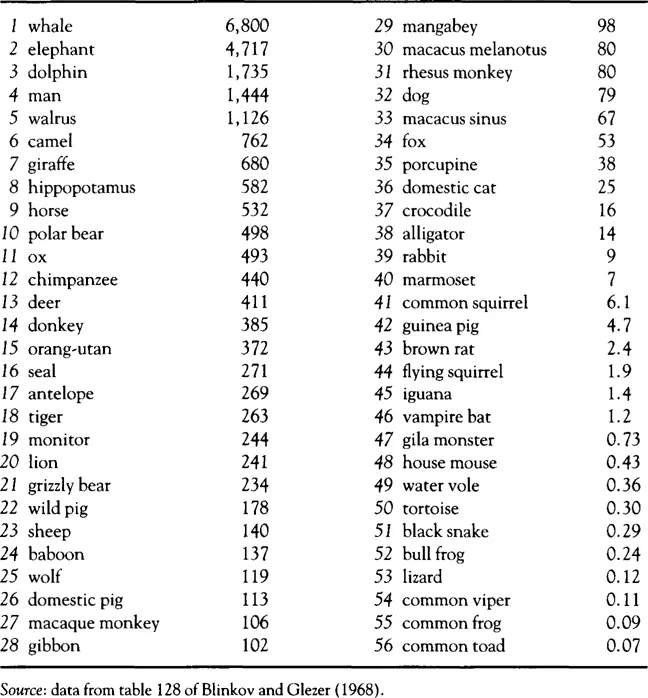
Absolute brain size (table 1.1) suggests a strong correlation between greater intelligence (or complexity of behavior) and absolute body weight, whereas this is apparently not the case either within a species or across species. Big animals are not necessarily smart ones.
Table 1.1 Brain sizes (in grams)
To eliminate the body-size factor, the ratio of brain size to body size has been examined, but conversely this index unduly favors various small animals (table 1.2) – with the flying squirrel and the devastatingly brilliant monitor finishing higher on this scale of intelligence than Homo sapiens. As seen in these tables, the spectrum of intelligence suggested by absolute brain weight may please those of us with a special dislike of snakes and frogs – and that suggested by brain/body ratios may offer some solace to those of us with a special fear of alligators, but neither measure gives man and the other primates their proper distinction.
The counter-intuitive nature of the hierarchies of intelligence in tables 1.1 and 1.2 has led to more complex coefficients of intelligence which are applicable only within specific limited groups (primates, mammals, etc.) (see Jerison, 1973, for a detailed discussion). From an evolutionary perspective, there are indeed grounds for arguing that rigorous comparisons will be possible only within such ‘lines’ of related organisms — and quantitative methods in comparative neuroanatomy currently focus on such subgroups. Yet without arguing against such techniques, it is nonetheless true that they cannot answer some of the questions which originally motivated research in comparative neuroanatomy. If we study evolutionary subgroups separately, how can we obtain an answer to questions about the relative intelligence of man, the dolphin and the songbirds? And can any conclusions be drawn about the human brain from assorted findings from comparative anatomy, such as the fact that the electric eel has a relatively large cerebellum or that the lion has a large amygdala?
Table 1.2 Brain weight/Body weight ratios
The relative intelligence of animal species may seem a prize topic for setting academics to sleep in their armchairs, but it is a part of a larger question of obvious importance. If behavior is controlled by (or at least through) the brain, then behavioral differences should reflect (or be reflected in) differences in brain function. When we are angry or fearful or attentive or confused, there simply must be characteristic brain states which are as consistent and replicable as are such cognitive states.
In the final analysis a complete ‘brain code’ would delineate what those differences are. At a more academic level we should be able to determine what, for example, the major neuronal differences are between the domestic dog and the wild wolf. Structurally their brains are very similar, but somewhere there are anatomical/physiological distinctions which make the dog amenable to domestic life and the wolf almost never so. What furthermore distinguishes the brains of schizophrenics from the brains of people capable of more or less normal social existence? Clearly we are yet a long way from explaining species differences, much less personality differences or psychiatric states in terms of brain structures, so that most research efforts have remained at the preliminary level of trying to correlate the crudest measures of personality, behavior or intelligence with the grossest measures of the brain itself.
In the study of animal species the ratio of cortical surface area to bulbar cross-section (a measure of the number of fibers ascending and descending through the brainstem) gets us closer to the expected hierarchy of intelligence, because we are then evaluating the degree of ‘excess’ brain matter beyond that needed for sensation and motor control of the body. The deficiency of this scale is that there is still a bias in favor of small animals. Ideally we would like to find a biological measure which is not affected by the size of the animal and which is applicable to a wide range of species.
Unfortunately no such measure has become established, but an interesting start has been made in Anthony’s (1938) ‘calloso-bulbar ratio’. As shown in figure 1.2, the calloso-bulbar ratio reflects the amount of nervous tissue devoted to communication between the left and right cerebral hemispheres, relative to the amount of tissue devoted to receiving/sending impulses up and down the spinal cord. Strictly speaking such an index is appropriate only for animals with a corpus callosum – ex...