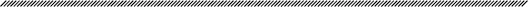![]()
1
Introduction to the Marine Algae
Overview
Ana Isabel Neto1,2,* and Isabel Sousa Pinto3,4
1 cE3c - Centre for Ecology, Evolution and Environmental Changes/Azorean Biodiversity Group, 9501–801 Ponta Delgada, Açores, Portugal.
2 Department of Biology, Faculty of Sciences and Technology, University of Azores, Rua da Mâe de Deus, 9500–321 Ponta Delgada, Açores, Portugal.
3 Interdisciplinary Centre of Marine and Environmental Research (CIIMAR), University of Porto, Terminal de Cruzeiros do Porto de Leixôes, Avenida General Norton de Matos, S/N, P-4450–208 Matosinhos, Portugal.
4 Department of Biology, Faculty of Sciences, University of Porto (FCUP), Rua do Campo Alegre, s/n, 4050 Porto, Portugal.
* Corresponding author
Definition
The term algae has no formal taxonomic standing but it is routinely used to indicate a large, polyphyletic, artificial, and diverse assemblage of mostly photosynthetic nonvascular organisms that contain chlorophyll a and have simple reproductive structures (Barsanti and Gualtieri 2014). As opposed to higher plants, algae do not have true roots, stems, leafy shoots, and transporting tissues; like mosses and ferns, algae do not form flowers and seeds, but reproduce and complete their life cycle by releasing gametes or spores into the environment. The algae concept encompasses a number of life forms that are not closely related; they are included in four kingdoms: Plantae, Chromista, Protozoa, and Bacteria (Ruggiero et al. 2015) including both prokaryotic and eukaryotic organisms (Lee 2008). The former, often classified as Cyanobacteria, are in fact photosynthetic bacteria, lacking the membrane-bounded organelles (plastids, mitochondria, nuclei, Golgi bodies, and flagella) that are present in the latter group.
The profound morphological diversity of forms and sizes, and the cytology, ecology, and reproductive biology of algae make it difficult to clearly define them (Bold and Wynne 1985). In a simplistic approach, the term algae refer to both macroalgae and a highly diversified group of microorganisms known as microalgae.
Species and environments
A conservative approach estimates the number of algal species in 72,500, most of them being microalgae (Guiry 2012).
Algae most commonly occur in water—marine, freshwater, and brackish—but they can also be found in almost every other environment on earth, for example, growing on snow in some high mountains and in the Arctic or living in lichen associations on bare rocks, in desert soils, and in hot springs (Lee 2008). In most habitats, algae are extremely important as they function as primary producers in the food chains and produce the oxygen necessary for their metabolism and that of the consumer organisms. Algae also form mutually beneficial partnerships with other organisms in which they provide oxygen and organic compounds and receive protection and nutrients. This is the case of the zooxanthellae that live inside the cells of reef-building corals, and of the green algae or cyanobacteria that associate with fungi to form lichens (Barsanti and Gualtieri 2014).
Algae cells contain the green pigment chlorophyll that captures the sun’s energy for photosynthesis, that is, the process of building energy-rich compounds from water and carbon dioxide. Photosynthetic algae will therefore grow only where there is light. In marine environments below low tide level, the amount and quality of light decreases with increasing depth to a point where the light level is not enough for algal growth. Some species contain additional pigments that enable them to absorb different light wavelengths and to use faint levels of light. This is the case of a coralline red algae collected at 268 m deep, the depth record for marine macrophytes (Littler et al. 1985). Due to these additional pigments, algae can exhibit different colors, the more common being red or brown. These shades are themselves extremely variable, with some brown species appearing olive-green and some red species almost black.
Many algae grow attached to the firm substratum (benthic). Benthic algae can grow attached on stones (epilithic), on sand or mud (epipelic), on animals (epizoic), or on other algae or plants (epiphytic). In marine environments, they can grow from the littoral zone (encompassing the intertidal area) to the shallow subtidal to around 200 m in very clear waters (Barsanti and Gualtieri 2014).
Most algae, however, are small single celled and filamentous organisms floating freely in the sea and constitute the phytoplankton, which forms the base of the marine food chains (Harris 1986; Sournia 2008). Most planktonic species are independent from the coastal and benthic processes, except for the temporary microscopic stages of the life cycle of the macroalgae (South and Whittick 1987; Falkowski and Knoll 2007).
Size and shapes
The size of algae is highly variable ranging from the tiny picoplankton which is only 0.2–2.0 μm in diameter to giant kelps with fronds up to 60 m in length.
Algae occur in dissimilar forms such as microscopic single cells (Fig. 1A), filaments (Fig. 1B), macroscopic multicellular loose or filmy conglomerations (Fig. 1C), colonies matted or branched (Fig. 1D), and more complex forms encompassing thin foliose sheets (Fig. 1E), tubes (Fig. 1F), sacs or bulbs (Fig. 1G), leather sheets (Fig. 1H), and gelatinous forms (Fig. 1I), cartilaginous (Fig. 1J) or calcareous forms, these including bushy (Fig. 1K), foliose (Fig. 1L) and crustose habits (Fig. 1M).
The unicells may or may not be solitary, and may or may not be motile through flagella. They can also exist as aggregates of single cells in colonial morphs, which can be more or less organized and have a variable number of cells. A colony is termed coenobium when the number and arrangement of cells are determined and remain constant. A good example of a motile coenobium is provided by the green alga Volvox (Fig. 2A).
The filamentous algae are formed by cells divisions along a plane perpendicular to the main axis in a way that all daughter cells are connected by their end wall. Filaments can be simple, as in Spirogyra (Fig. 2B), or branched. Branching can be further classified as false (as in the Cyanobacteria Tolypothrix,Fig. 2C) or true (as in the green Cladophora, Fig. 2D). Filaments can also consist of a single layer of cells (uniseriate) as in the red Lejolisia (Fig. 2E) or made up of multiple layers (multiseriate) as in Polysiphonia (Fig. 2F).
A peculiar form occurs in the siphonous algae, which have a coenocytic construction, consisting of tubular filaments that grow as their nuclei undergo repeated nuclear divisions but in which transverse cell walls never form. The resulting filament is therefore unicellular but multinucleate (coenocytic). A classic example of a branched coenocyte thallus is found on the green alga Bryopsis (Fig. 2G).
Fig. 1. Examples of algae with different shapes: (A) unicellular as in Haematococcus, (B) filamentous, (C) cyanophyceae film, (D) colonies as in Haematococcuspluvialis, (E) thin foliose sheets as in Porphya, (F) foliose tubes as in Ulva intestinalis, (G) bulbs as in Colpomenia sinuosa, (H) leader sheets as in Laminaria, (I) Gelatinous, (J) cartilageneous as in Cystoseira abiesmarina, (K) calcareous bushy as in Ellisolandia elongata, (L) calcareous foliose as in Padina pavonica, and (M) crustose habits as in Peyssonnelia squamaria. Photos from Island Aquatic Research, Azorean Biodiversity Group/Center for Ecology, Evolution and Environmental Changes (IAE/GBA/cE3c).
Fig. 2. Morphology of algae (A) motile coenobium as in Volvox, (B) filamentous simple, as in Spirogyra, (C) filamentous with false branches as in the Cyanobacteria Tolypothrix, (D) filamentous with true branches as in Cladophora, (E) uniseriate filaments as in the red Lejolisia, (F) multiple layers (multiseriate) as in Polysiphonia. Photos from Island Aquatic Research, Azorean Biodiversity Group/Center for Ecology, Evolution, and Environmental Changes (IAE/GBA/cE3c).
The more complex types and forms (cartilaginous) have a parenchymatous or pseudoparenchymatous construction, which originates from a meristem with cell divisions in three dimensions. In the parenchymatous thallus, cells of the primary filament divide in all directions and the essential filamentous structure is lost (e.g., Ulva, Fig. 2H). The pseudoparenchymatous thallus is made by the aggregation, in a loose or compact way, of numerous, intertwined, branched filaments held together by mucilages to collectively form the thallus, which therefore has little internal differentiation. A good example is found in the red alga Schizymenia (Fig. 2I).
Reproduction
The reproduction in algae is highly variable. Asexual reproduction, arguably the most widespread, can be vegetative (by the division of a single cell or by the fragmentation of a colony or filament) or by the production of motile spores. This mode of reproduction allows stability of an adapted genotype from one generation to the next, but restricts genetic variability. Sexual reproduction, involving the union of gametes, results in the genetic recombination needed for adaptation to changing environments (Lee 2008).
The binary fission of unicellular algae is the simplest form of asexual reproduction, in which the parent organism divides into two equal parts, each containing the same hereditary information as the parent. The autocolony formation is a specific mode in which each cell of the colony can produce a new colony similar to the one to which it belongs. Cell divisions produce a sort of embryonic colony smaller than the parent colony but equal in cell number (Fig. 2A).
In the fragmentation mode, the filaments and/or non-coenobic colonies break into two or several parts, each having the capacity of developing into new individuals.
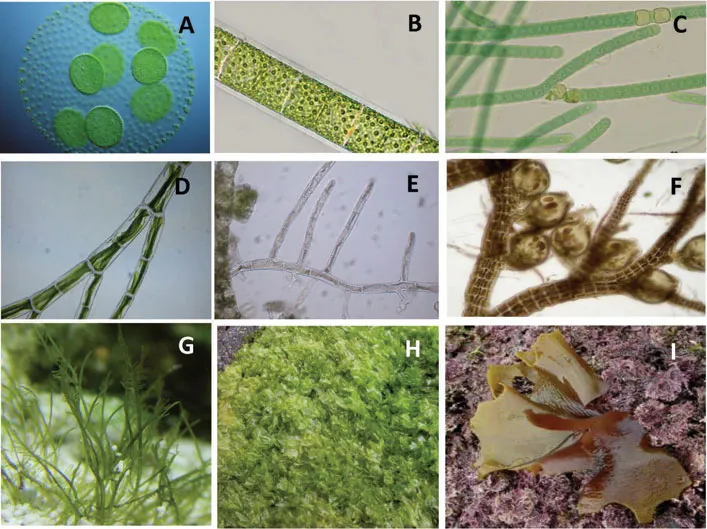
Asexual spores can be flagellate motile (zoospores) and produced by a parental vegetative cell, aflagellate (aplanospores) that begin their development within the parent cell before being released, and autospores that are aflagellate daughter cells that will be released by the rupture of the parent cell wall. These later are almost perfect replicas of the mother cell as in the green alga Chlorella (Fig. 3). Spores may be produced within ordinary vegetative cells or within specialized structures called sporangia.
Fig. 3. Chlorella spores resembling mother cells. Photos from Island Aquatic Research, Azorean Biodiversity Group/Center for Ecology, Evolution and Environmental Changes (IAE/GBA/cE3c).
Under unfavorable conditions many algae can produce thick-walled resting cells, such as hypnospores, hypnozygotes, statospores, and akinetes, and enter a dormancy period. The former two are produced by protoplasts that were previously separated from the walls of the parental cells. They enable the species to survive temporary drying out of water bodies. Statospores are endogenous cysts formed within the vegetative cell. Their silica enriched walls, often ornamented with spines and other projections, make them good fossil records. Akinetes are of widespread abundance in the blue-green algae and used in Alfa taxonomy. They are essentially enlarged vegetative cells that develop a thickened wall in response to limiting environmental nutrients or limiting light.
The various types of gametes can be classified according to relative size and motility. When both are motile and the same size (isogametes) the process is called isogamy; when gametes differ in size (heterogametes) they can undertake the anisogamy process, when both are motile but one is small (male, by convention) and the other is large (female); or the oogamy process, in which only the male gamete is motile and fuses with a very large and non-motile female gamete.
Gametes may also be morphologically identical or markedly different from vegetative cells, depending on the algal group. When similar they can only be distinguished based on the DNA ploidy.
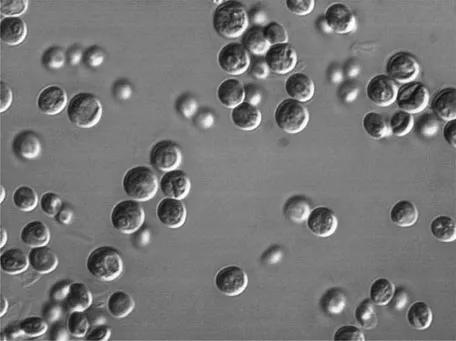
The meiosis occurrence in a sexual life cycle and the type of cells it produces determines different types of reproductive cycles. If the meiosis occurs immediately after the zygote formation and the cycle is predominantly haploid it is called the haplontic or zygotic cycle (Fig. 4). If the cycle is predominantly diploid and the meiosis give rise to haploid gametes that are short lived and fuse to form a new zygote, the cycle is called diplontic or gametic (Fig. 4). On the diplohaplontic or sporic life cycle (Fig. 4) there is an alternation of generations between two phases, one haploid, represented by the gametophyte, and the other diploid, represented by the sporophyte. The former produces gametes by mitosis, the later produces spores through meiosis. If the two generations are morphologically identical, the alternation of generations is isomorphic. If they are different, the alternation of generations is hetermorphic.
Fig. 4. Adapted from 20 Bell, G. 1994. The comparative biology of the alternation of generations: Lectures on mathematics in the life sciences: Theories for the evolution of haploid–diploid Life Cycles. 25: 1–26.
Evolution and classification
Evolution
Biotic interactions between algae and other eukaryotes are very common in aquatic and terrestrial ecosystems. These interactions can be intracellular and extracellular/surface interactions in the phycosphere, which is the ecologically and physiologically integrated neighborhood inhabited by the alga (Brodie et al. 2017). Epibiosis (surface colonization of one organism, by other organisms, epibionts) occurs on all immersed surfaces, including those of micro- and macroalgae. Epibiotic interactions (e.g., alga-alga, alga-bacterium, alga-virus) play key roles in nutrient acquisition and recycling, metabolic flux, energy flow, and developmental processes. Together with herbivory, epibiosis represents one of the most important interactions for algae and has been shown to shape entire marine communities (Brodie et al. 2017).
A very important contribution to our knowledge of the evolution of life on earth was the endosymbiotic theory by Lynn Margulis (1981) according to which s...