Originally published in 2002, Mountain Biodiversity deals with the biological richness, function and change of mountain environments. The book was birthed from the first global conference on mountain biodiversity and was a contribution to the International Year of Mountains in 2002. The book examines biological diversity as essential for the integrity of mountain ecosystems and argues that this dependency is likely to increase as environmental climates and social conditions change. This book seeks to examine the biological riches of all major mountain ranges, from around the world and using existing knowledge on mountain biodiversity, examines a broad range of research in diversity, including that of plants, animals, human and bacterial diversity. The book also examines climate change and mountain biodiversity as well as land use and conservation.

- 346 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
EcologyIndex
Biological SciencesPart I
Introduction
1
Mountain Biodiversity, its Causes and Function: an Overview
The high elevation biota of the world have been the subject of many synthetic approaches, including assessments of the variability and diversity of ecosystem properties and processes, ecophysiological traits and organismic functioning in general (e.g. Franz, 1979; Vuilleumier and Monasterio, 1986; Billings, 1988; Rundel et al., 1994; Archibold, 1995; Wielgolaski, 1997; Körner, 1999). In this overview, questions relating to high elevation biodiversity will be specifically discussed (for previous attempts with a global perspective see for instance Chapin and Körner, 1995; Rahbek, 1995).
Nails and Screws and the Lowland-Upland Contract of Society
Mountainous terrain covers 24% of the global land area (Kapos et al., 2000). The trivial fact that mountains have slopes causes them to directly or indirectly influence the life of half of the Earth’s human population (Messerli and Ives, 1997). Slopes (and the peaks behind them) not only capture water, but guide it to the forelands and, via large river systems, feed the plains. Runoff and the sediments carried with it, both benefit (water supply and mineral nutrients) and disadvantage (floods and mud slides) downslope society. Slopes provide gravitational power to water, which can be converted to electric energy. Slopes guard and guide or constrain and endanger traffic routes. Slopes stop clouds (advection) or create them (convection), slopes exert mechanical force on any organism, installation or activity on them.
Unless made of solid rock, the only way in which loose substrates are secured to slopes is through vegetation. These living claws, screws and nails sustain slope benefits and prevent slope dangers. Slopes are only as stable and safe as the integrity and stability of their vegetation (Figure 1.1). Integrity of upslope vegetation is the centrepiece of downslope welfare in many parts of the world. There is, indeed, a too often overlooked link between far away mountains and densely populated lowlands. The recurring floods in China, for instance, which directly affected 200 million people’s lives in 1999, the recent mud slides in the Philippines or the Andes, are impressive manifestations of uncontrolled upslope instability. It is obvious that there are many mutual upslope-downslope interdependencies. A substantial fraction of lowland productivity is required to cover the needs of upslope society and to maintain or enforce sustainable management in the highlands. This ‘downslopeupslope contract’, in essence, rests on the effectiveness of vegetation - natural or man-made - to control erosion.
Biodiversity Provides Insurance
The formula is simple: intact vegetation provides safety. Since the risks and threats on mountain slopes are manifold, they require a multitude of safeguards. The hammering of the ground by heavy raindrops or hail, the disruptive force of surface runoff, the mechanical sheer of loose ground, the exploration of deep substrate moisture, the resistance against trampling and grazing by large herbivores, and snow-gliding, are among many manifestations that require different mechanical solutions (see Figure 1.1). A multitude of plant structures can and do offer these services in a concerted manner. Yet, at times, any of these ‘tools’ may fail. Natural diseases, divergent life cycles, and varying sensitivity to stress and disturbance may eliminate different players, at least periodically. Hence, it requires each key tool to be present in various combinations, so that at least one functions in case of emergency. The benefits of diversity are manifold (Mooney et al., 1995). In the simplest terms, Moffat, (as quoted by Stuart Pimm 1996) said that: ‘There is a dance going on of compensatory changes. Something always benefits from a disaster, provided you have enough species’. Whether ecosystems with more species are less at risk of loss of integrity than systems with less species is not a generalizable issue, as was previously thought. It depends on the absolute number of species, the identity of the species and the circumstances under which the system is operating. Hence, the above discussion should be taken as a plausible trend, not a proven fact for any individual case.

Figure 1.1 The integrity of steep alpine slopes such as this one, at 2500 m in the Swiss central Alps, is secured by plants, their belowground structures in particular. The cartoon to the right symbolises the need for a multitude of ‘tools’ to do the ‘job’
According to the ‘insurance hypothesis’, one of the benefits of biological richness is that it insures against system failure. Functional redundancy may not play any role for long periods of time, but a single extreme event can cause it to become the life-preserver of fragile slopes that sustain ecosystem functioning. The richness of biota includes all levels of organismic diversity from genes to landscapes, it includes all groups of organisms from microbes to the largest herbivores.
By no means should ecosystem functioning be considered the only valid reason to conserve biological richness, but prioritising on this one function appears plausible, given that all other benefits and values depend on it. Other justifications are the ethical value (the right of existence), the aesthetic value (the beauty), the economic value (potential or instrumental economic uses). Each of these has value in its own right. In high mountains, several or all of these values apply together, as this book will illustrate.
Having said all this, the reader will find this introductory note somewhat plant-biased. The reason for this is, in part, my own profession. However, having joined many discussions with zoologists, there seems to be less synthetic knowledge available in high elevation animal sciences. Perhaps we are far from even simple generalisations which may hold for animal studies. As an example, plants become smaller with increasing elevation, if the same morpho-types are compared; not so animals: birds and rodents become larger, as do some groups of insects, others do not significantly change in size and some also become smaller (such as fish). If they become smaller, are there more individuals needed to fulfil a particular ecosystem function (such as chopping down litter)? It seems, we do not know. Do developmental cycles become shorter? For grasshoppers, this does not seem to be the case. Thankfully a few animal scientists have joined this assessment and the doors are wide open for more to come in!
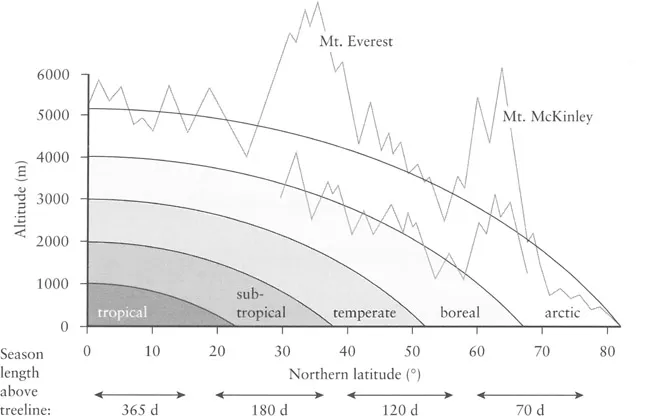
Figure 1.2 The correlation between altitudinal and latitudinal life zones. The elevational compression of biomes causes mountains to become hot spots of biological richness
The Biological Richness of High Elevation Biota
As one ascents a humid, 5000-m high equatorial mountain, one passes through nearly all climatic zones of the world over a relatively short distance (Figure 1.2). On flat land, one would have to travel thousands of kilometres to pass through a similar series of thermal climates. Imagine a bear in the Rocky Mountains or the Carpathians, on a pleasant autumn afternoon it may be the berries in the alpine heathland which attract its attention, while the ‘boreal’ forest below provides shelter for it during the following cold night. Heavy snowfall may lead the bear to spend the next morning in the deciduous forests of the temperate life zone a few kilometres downhill. Within a day, the bear has covered a 3000-km range of latitudinal climatic difference, while perhaps walking no more than 8 km. This compression of life zones explains why, on a 100 km grid scale, no landscape can beat the biological richness of mountains (Barthlott et al., 1996). Nowhere else is it possible to protect and conserve so much biological diversity within a relatively restricted region, than in mountains and in the tropical mountains in particular (Klötzli, 1997). Tropical mountains may span the range from the humid lowland forest to glaciated mountain tops, and it is therefore no surprise that these regions become hot spots of biodiversity.
The richness of organismic taxa declines with elevation (at least in humid areas), and biologists have long recognised that elevational and latitudinal species richness gradients mirror each other (Figure 1.2; Rahbek, 1995 and references therein). Once the glacial region is reached, it is unlikely that more than 5% of all species of a given mountain region and its immediate lowland surroundings can thrive. Within the alpine belt, the total plant species diversity of a given region commonly declines by about 40 species of vascular plants per 100 m of elevation (Figure 1.3). Surprisingly, this rate of decline does not differ greatly across a wide range of mountains (see Nagy et al., in prep.). This decrease in species richness follows the decrease in mean temperature (Figure 1.4), but often accelerates in the upper part of such transects.
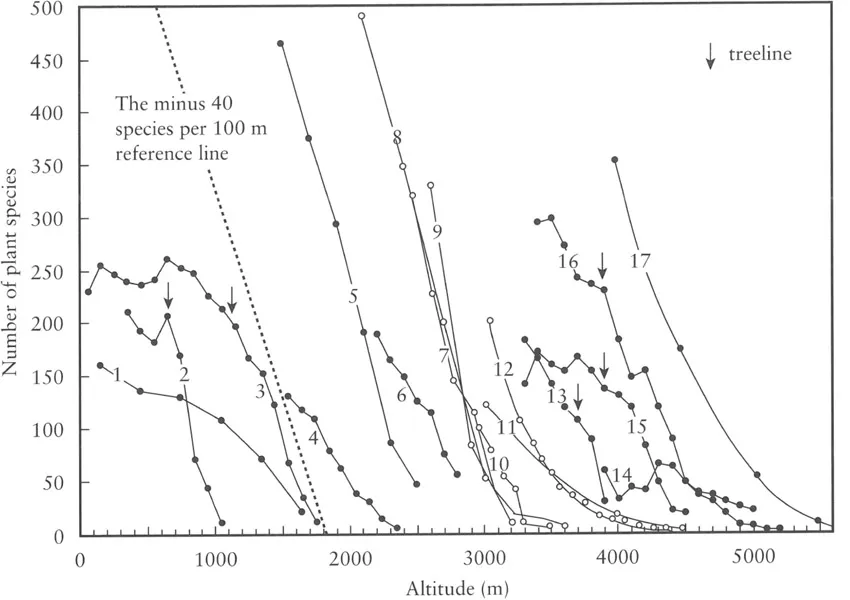
Figure 1.3 Examples of the elevational reduction of plant species diversity in different mountain ranges. (1) E, NE Greenland (Schwarzenbach, 2000); (2) Glen Clova, Scotland (Willis and Burkill in Raunkiaer, 1908); (3) Aurland, S Norway (Odland and Birks, 1999); (4) Jotunheimen, S-Norway (Jorgensen, 1932 in Odland and Birks, 1999); (5) Tatra Alps (Kotula in Raunkiaer, 1908); (6) Olympus, Greece (Polunin, 1980); (7) Swiss Alps (Heer in Raunkiaer, 1908); (8) Bernina, Swiss Alps (Brockmann-Jerosch in Raunkiaer, 1908); (9) West Alps (Thompson in Raunkiaer, 1908); (10) Bernina, Swiss Alps (Rübel, 1911); (11) Oetztaler Alps, Tyrol (Reisigl and Pitschmann, 1958); (12) Montafon, Alps (Grabherr et al., 1995); (13) Oytagh; (14) K2 North; (15) Batura; (16) Nanga Parbat, Karakorum (Dickoré and Miehe, Chapter 10); (17) Hindukush (Breckle in Walter and Breckle, 1991)
Species richness in animals declines with elevation in a similar way as in plants. Rahbek (1995) reviewed this field most recently and, from his analysis of 97 publications with 163 examples, it is quite obvious that altitude and latitude operate in the same direction. However, Rahbek noted that this decline is not necessarily monotonie (Figure 1.5), and Meyer and Thaler (1995) showed that trends strongly depend on the taxonomic group considered. The reasons for this elevational decline of species richness are not fully resolved. Since similar trends are observed in plant and animal diversity, we are either dealing with a general evolutionary bottleneck phenomenon (for whatever reason), affecting all organismic groups similarly but independently, or the reduced plant diversity induces the decline in diversity of all other organismic categories. Cryptogams (mosses and lichens) may deviate from this pattern over certain ranges of elevation and increase in abundance as the climate becomes cooler and more humid (Virtanen et al., 2002), but ultimately the number of cryptogam species also drops drastically as one reaches the highest peaks. Isolated individuals of higher plants have been found up to 5900 m in Tibet, 6300 m in the Himalayas, and up to 4450 m in the Alps (Rongfu and Miehe, 1988; Grabherr et al., 1995; Grabherr, 1997; Körner, 2001), but these outposts ofte...
Table of contents
- Cover
- Half Title
- Title
- Copyright
- Original Title
- Original Copyright
- Contents
- Preface
- Part I: Introduction
- Part II: How much mountain biodiversity is there and why?
- Part III: Climatic changes and mountain biodiversity
- Part IV: Mountain biodiversity, land use and conservation
- Part V: Synthesis
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Mountain Biodiversity by Ch. Korner,E.M. Spehn in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Ecology. We have over 1.5 million books available in our catalogue for you to explore.