
- 452 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
The 1985 Research Conference on Fire Ants and Leaf-Cutting Ants covered the most recent developments in research and control of these insect pests of the New World tropical and subtropical zones, the southern United States, South and Central America, and the Caribbean Islands. This volume contains chapters that discuss the history and economics, biology and ecology, behavior, pheromones and other semiochemicals, physiology, and biochemistry of fire ants and leaf-cutting ants, as well as current and future control strategies. The information provided illustrates past and present agricultural and medical problems associated with these pest ants; however, it also brings out the point that they may at times be beneficial. The chapters on basic aspects of the biology and ecology of the ants provide up-to-date information that is useful for a more complete understanding of their social behavior and the unique symbiotic relationship between leaf-cutting ants and their fungi. New approaches to control are illustrated by innovative research on anti-feedant chemicals from plants that prevent feeding by leaf-cutting ants. The present status of chemical baits and biocontrol is addressed, as well as the possibilities of future novel methods based on the use of anti-metabolites, insect hormones, behavior modifying substances, and species-specific toxic bait systems to create integrated pest management systems.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
BiologyIndex
Biological SciencesBiology and Ecology
7
The Ecological Nature of the Fire Ant: Some Aspects of Colony Function and Some Unanswered Questions
W. R. Tschinkel
Presented here is a conceptual interpretation of the available facts about fire ants that will be used to paint a general picture of their biological nature and function. Considering the gaps in our knowledge, the future may very well see changes of interpretation. The facts upon which I base my review are the results of the work of many people over more than three decades. Information on history and spread of Solenopsis invicta is discussed by Lofgren in Chapter 4.
Ecological Nature of S. Invicta
Ecologically, S. invicta is a weed species and, as such, shows many of the biological properties of weeds. Weeds are animal or plant species that are adapted for the opportunistic exploitation of ecologically disturbed habitats. Naturally, these are created by flood, fire, and storm and consist of new sandbanks, slumps, and landslides, burns, and windfalls. Man, however, creates vast areas of disturbed habitat by clearing forest for agricultural, domestic, and other uses. The plant and animal communities that occupy such disturbed habitats are called early secondary or early succession communities because, if left alone, they will gradually revert to dominant climax communities. In the southeast, this is mostly deciduous forests. Because such early succession communities are ephemeral and underexploited, the weed species utilizing these habitats are adapted for very rapid, scramble-type exploitation with an emphasis on high reproductive rates and efficient dispersal rather than competition with other members of the community (Ito 1980).
The weed-like properties of the fire ant are as follows: First, the fire ant is clearly and dramatically associated with ecologically disturbed habitats created mostly by man both in the United States and South America (Banks et al. 1985). Thus, S. invicta is abundant in old fields, pastures, lawns, roadsides, and any other open, sunny habitats. It shares these habitats with many other weedy plant and animal species, from man's crops to lawn and pasture grasses, goldenrods, and dog-fennel. Man is the fire ant's greatest friend, even though the sentiment may not be returned.
On the other hand, the fire ant is absent or rare in late succession or climax communities such as mature deciduous or pine forest (personal observation). When it is found in these communities, it is usually associated with local disturbances such as seasonal flooding and roads. In North Florida, on transects through longleaf pine-wiregrass-turkey oak forest, I found S. invicta strictly associated with temporary ponds or pond margins, graded dirt roads, and the margins of paved roads. All other areas were occupied by S. geminata, if any fire ants were present at all. This was true even in recently clearcut and replanted areas. Increased insolation associated with disturbance is thus not sufficient explanation for S. invicta's distribution. Unfortunately, there are almost no data on the biotic and abiotic causes of its distribution. Nevertheless, it is clear that S. invicta, like other weeds, is associated with open, disturbed habitats. This also appears to be true in its native homeland in Southern Brazil (Buren, personal communication; Banks et al. 1985).
A high reproductive rate is a second weedy property that is associated with the sporadic, unpredictable and ephemeral availability of suitable habitat (in the absence of man-made disturbance). Success in such a situation goes to the animals and plants that "gits thar the fustest with the mostest" (attributed to Civil War General Lee DeForrest in response to being asked how to win a battle) with little attention paid to competition within the community. Fire ants, like other weeds, achieve a high reproductive rate in part by very high investment of resources in reproductives. From the meager data available (Markin et al. 1973; Morrill 1974), I estimate that S. invicta allocates 30 to 40% of its annual biomass production to sexuals. This is similar to the energy allocation to seeds found in weedy species of goldenrod and much higher than non-weedy goldenrods adapted to competition in late-succession communities (Ito 1980) (Fig. 1). Thus, the average fire ant colony in North Florida produces about 4500 sexuals per year (Morrill 1974). Although very little information is available for comparison, this seems high for ants in general and is almost certainly an adaptation to its weedy habits.
A third weedy property is effectiveness of dispersal and colonization. In the absence of man-made disturbance, secondary habitat is scattered and unpredictable. Its exploitation depends upon the ability to scatter propagules (sexuals or seeds) over wide areas on the chance that a few will find their way to an appropriate site and colonize it. The fire ant achieves this by producing a large number of sexuals who take part in high-altitude, dispersive mating flights throughout a large portion of the year (Fig. 2). The queens often fly or are wind-carried one-fourth to one-half mile or more before settling to the ground although most settle at shorter distances (Markin et al. 1971).
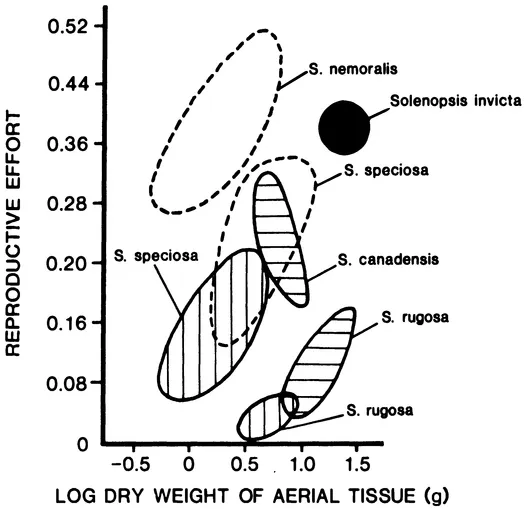
FIGURE 1. Reproductive effort (proportion of production invested in seeds or sexuals) in relation to size of organism or colony for goldenrods (Solidago spp.) and fire ants (black circle). Each enclosed area is represented by the individuals of a single population on a dry site (enclosed in dotted curve), a wet site (with horizontal shading), or a hardwood site (vertical shading) (modified from Ito 1980).
Fire ant queens do not have to depend on general habitat disturbances to enhance their ability to establish a colony; even a specific disturbance to the ant community can provide the necessary conditions for success. This phenomenon was first observed by Summerlin et al. (1977) in mirex-treated plots in which S. invicta was a minor component of the ant community. The mirex killed almost all of the ground-nesting ants; but after recolonization, S. invicta and another weedy species, Conomyrma insana, had greatly increased their dominance over all other species (Fig. 3). Many of the native species did not reappear in the course of this two-year study.
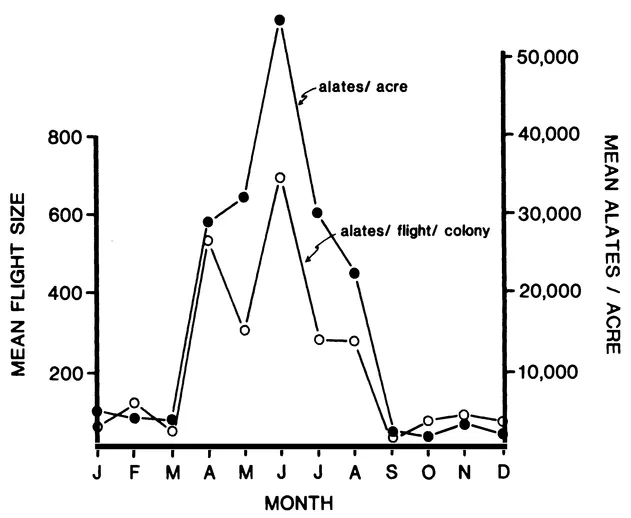
FIGURE 2. Occurrence and size of mating flights throughout the year in North Florida (from Morrill 1974).
The implication of these studies is clear: Large-scale, unspecific control programs utilizing chemical baits in areas of low fire ant populations may aid rather than hinder the estab...
Table of contents
- Cover
- Half Title
- Title
- Copyright
- Contents
- Foreword
- Preface
- Acknowledgments
- KEYNOTE ADDRESS
- HISTORY AND ECONOMICS
- BIOLOGY AND ECOLOGY
- PHYSIOLOGY AND BIOCHEMISTRY
- CURRENT AND FUTURE CONTROL STRATEGIES
- List of Contributors
- Taxonomic Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Fire Ants And Leaf-cutting Ants by Clifford S. Lofgren in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.