
- 154 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
This study of plant anatomy is based on newly available data on the structure and spatial organization of the vascular system of plants. For the first time, by means of a new technique of intracellular moulding, the vascular system can be observed in its length. Many examples are chosen from among the major groups of the plant kingdom to illustrate
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
BiologyIndex
Biological SciencesPart I
Potential Applications of Microcasting
Histogenesis, Structure and Function of the Xylem
OVERVIEW
The cells of terrestrial plants are differentiated1 into tissues, each with differing morphology and function. Differentiation is absent or weak in Thallophytes and is accentuated in the higher evolutionary plant forms to reach its most advanced stage in the Angiosperms (Table 1). With differentiation, the vital functions of the plant have been progressively allocated to different tissues and, consequently, exchanges of matter and signals have been developed between its various parts. The capture of solar energy and the fixing of atmospheric carbon, for example, have become the specific functions of the photosynthetic tissue and the foliar mesophyll, the absorption of water and dissolved minerals from the soil has become that of the young root tissues at the other end of the plant, and the distribution of absorbed and metabolized substances is that of the vascular tissues, the xylem and phloem, in the species endowed with those tissues, the Tracheophytes.
It has been at least three centuries since the circulation of water and matter in the plant was foreseen from simple observations (Grew, 1682; Malpighi, 1686) and about a century and a half since the vascular tissues were located and named (Hartig, 1837, 1854; Naegeli, 1858; Sanio, 1873, 1874). Research has progressed steadily since, and histogenesis2 and the structure and functions of the xylem and phloem continue to be fascinating aspects of the study of plant anatomy and physiology.
The xylem and phloem, which were named by the botanist Naegeli, are the pathways of circulation of two flows of matter. The greater flow in terms of mass is the hydric flow of raw sap through the xylem, rising from the soil and supplying all the tissues; most of the water is transpired by leaves and evaporates into the atmosphere. The latter flow, slower and slighter in volume, is the transport of concentrated solutions of carbon metabolites produced by photosynthesis to growing and reserve organs via the phloem.
Table 1. Classification of the plant kingdom
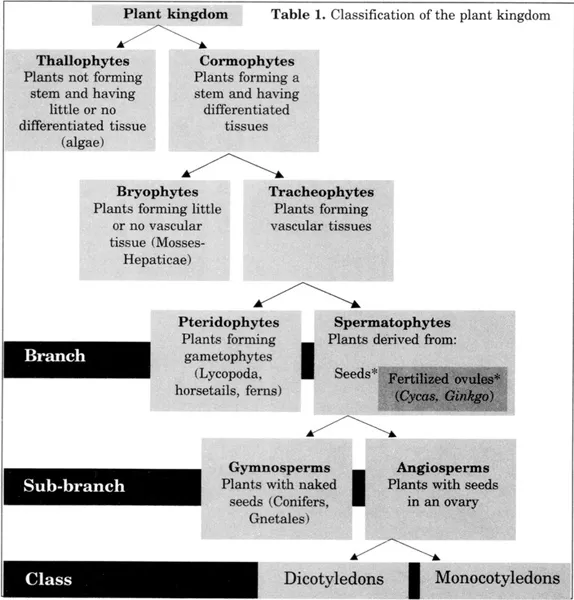
* The seed is a fertilized ovule that maintains exchanges with the mother plant until maturity. A parallel has been established with ovipary and vivipary in the Animal Kingdom (Emberger, 1960).
The two tissues form two closely associated networks, which extend as the plant grows and which draw its architecture with the finest details from the roots to the leaves, flowers, and fruits. Their histogenesis explains their perfect superposition at every point: their cells are in fact derived from divisions of the same meristem, the cambial meristem,3 which stretches between them like a thin tissue.
In this book, which is devoted to certain aspects of the morphology and spatial organization of the xylem, the presence and role of the phloem are mentioned only incidentally. The overall concepts of histogenesis and histology of the xylem are reviewed in this section so that they can be used to relate the activity of the cambial meristem to xylem structures described in the following sections.
HISTOGENESIS, STRUCTURE AND FUNCTION OF THE XYLEM
The histogenesis of conducting tissues and of the xylem in particular is a difficult process to capture in its spatio-temporal dynamics. To give a brief description, at the tissue scale to start with, let us follow the growth of a shoot of a ligneous Dicotyledon from an apical bud of the preceding year (Fig. 1.1A). From its apex to its lower part, this shoot will comprise at any given moment all the stages of xylem formation.
• Primary and secondary xylem of dicotyledons
At stage 1 of its growth, all the cells of the shoot grow in length. The procambial meristem, partly preformed in the bud, extends the interconnected network of its strands up to the apex. The primary xylem and phloem are differentiated from these strands, the xylem from their inner cells and the phloem from their outer cells (Fig. 1.1, section S1). The bundles, formed of these three tissues, are extended into the leaf petioles.
At stage 2, most of the basal cells of the shoot have completed their elongation, while the shoot continues to lengthen by division and growth of the younger cells. The meristem is now composed of a basal part, the cambium, and an elongating distal part formed of strands of the procambium. While the latter continue to produce primary xylem and phloem, the cambium laterally extends into the interfascicular regions and forms a double sheath of secondary xylem and phloem. After the elongation, the shoot grows in diameter (Fig. 1.1, section S2). The secondary tissues extend towards the apex (stage 3) until they reach the base of the new bud, when the growth of the entire organ is completed (stage 4).
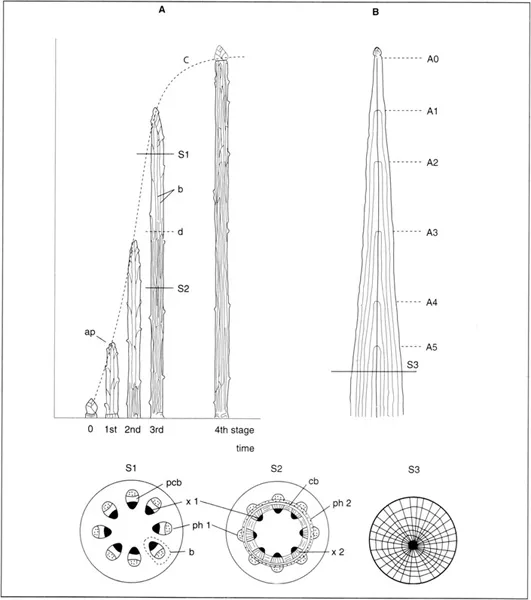
Fig. 1.1. Shoot growth in a ligneous Dicotyledon. A. Successive growth stages between the apical bud of the branch (stage 0) and the end of growth (stage 4). (On the diagrams, the leaves are sectioned at the base of the petioles. The shoot is shown with bark stripped.) The longitudinal growth (curve C) results from the cumulative elongation of all the cells after their formation by division close to the apex (ap). The spaced-out vertical lines at stages 1, 2, and 3 represent the primary bundles (b) in the growing part of the shoot; the horizontal line d delimits the basal part of stage 3, in which the growth is completed: the primary xylem (x1) is progressively ensheathed from the base to the apex by the secondary xylem (x2), represented by short lines close together. The lower part of the figure shows two cross-sections (SI and S2) respectively above and below the limit d of the two parts: only the meristematic continuum (procambium pcb + cambium cb) and the primary and secondary xylem (x) and phloem (ph) are illustrated. B. A branch is composed of units of successive growth. Seen in axial section, each annual deposit of wood covers and extends the preceding one; seen in cross-section (S3), it forms a supplementary ring (A0, A1, etc.: years of growth).
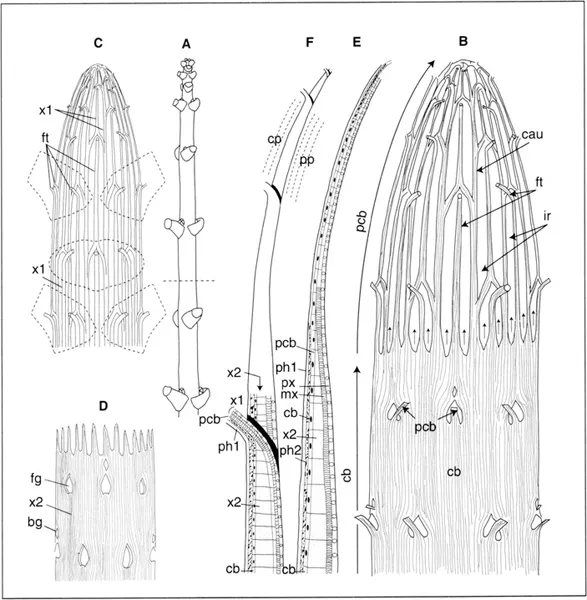
Fig. 1.2. Arrangement of the meristematic procambium-cambium (pcb-cb) continuum and the primary and secondary xylem (x1, x2) in a maple shoot; A. Diagram of the shoot in which the leaves are sectioned. The dottted line marks the present lower limit of the part that is still growing and thus the upper limit of the cambium and wood. In diagrams B, C, and D, the diameter-length ratio of the shoot is dilated by a factor of 8. B. Procambium-cambium continuum: the procambial network produces cauline bundles (cau) the branches of twhich form foliar traces (ft). Lower down, the interfascicular regions (ir) are progressively occupied by the extension of the cambium (arrows); C. The network of the primary xylem (x1), copy of the pcb network, extends from the base to the apex of the shoot; D. The wood or secondary xylem (x2), produced by the cb, progressively ensheaths x1, except at the points of emergence of the primary xylem of leaves (and of non-represented buds) at the “gaps” (foliar gap and bud gap, fg and bg); E. Diagram, in radial section along a cauline bundle, of the superposition of meristems (pcb, cb), primary and secondary phloem (phi, ph2), primary xylem (x1), itself made up of protoxylem, then metaxylem (px, mx)...
Table of contents
- Cover
- Half Title
- Title Page
- Copyright Page
- Table of Contents
- Dedication
- Foreword
- Preface and Acknowledgements
- Introduction
- I Potential Applications of Microcasting
- II The Technique of Microcasting
- References
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Vascular Organization of Angiosperms by Jean-Pierre Andre in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.