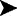1.1.1. The range of living organisms and the place of plants
a) The major divisions of the living world
Life is closely linked to the chemistry of carbon and water. Sugars or glucides, essential components of living matter, have long been considered hydrates of carbon. Lipids are made up of three basic elements: carbon, hydrogen, and oxygen. Only the proteins have a fourth element—nitrogen, which plays an important role in the diversity of living organisms. All these elements also belong to the inert mineral world, solid or gaseous, and are constantly involved in exchanges between that inert world and the living world. These exchanges and matter flows, which consume a large amount of energy, constitute the major cycles of water, carbon, and nitrogen. “Life” seems to be the essential driver of such periodic cycling of elements.
The extraordinary diversity of living forms has necessitated the conception of a classification of these forms, putting into a single group those individuals that share a certain number of common characteristics. This is how, for example, the prokaryotes, which like bacteria do not have a nuclear compartment, are distinguished from eukaryotes, which carry most of their information in a territory called the nucleus. Eukaryotes are themselves subdivided into plants and animals according to whether or not they are equipped and able to use light energy. A preliminary approach to the classification of living organisms is thus based on three major, clearly distinguished groups. There are also intermediate groups, often constituting interesting transitions that merit a closer look.
b) Definition of a plant
Generally, a plant is defined by its capacity to use light energy, i.e., energy linked to light radiation, to combine water molecules and carbon atoms to synthesize sugars. The sugars constitute energy reserves that allow plant cells to carry out all the other syntheses necessary for their metabolism. This carbon-based autotrophy of plants is based on the presence of specific pigments such as chlorophylls, while animals, which do not have this capacity for synthesis, use plant sugars and are thus entirely dependent on plants. They are situated further along the food chain. This essential criterion, however, is not the only one and normally the following criteria are added, in no particular order.
the existence of meristems, which give plants a constant and indefinite growth;
a general totipotency of cells that preserves the capacity of regeneration throughout the life of the plant, i.e., the ability to recreate a new plant;
meiosis, which results in the differentiation of spores and not gametes;
an alternation of generations, gametophyte and sporophyte, separated by fertilization and meiosis;
the presence of rigid framework of cellulose around each cell;
the existence of a triple heredity—nuclear, plastidial, and mitochondrial;
a kinetic apparatus that has some variability in its organization, for example, centrioles replaced by polar crowns;
male gametes that do not always have the capacity for autonomous displacement;
a fixed life.
c) Frontiers of the plant kingdom
With prokaryotes
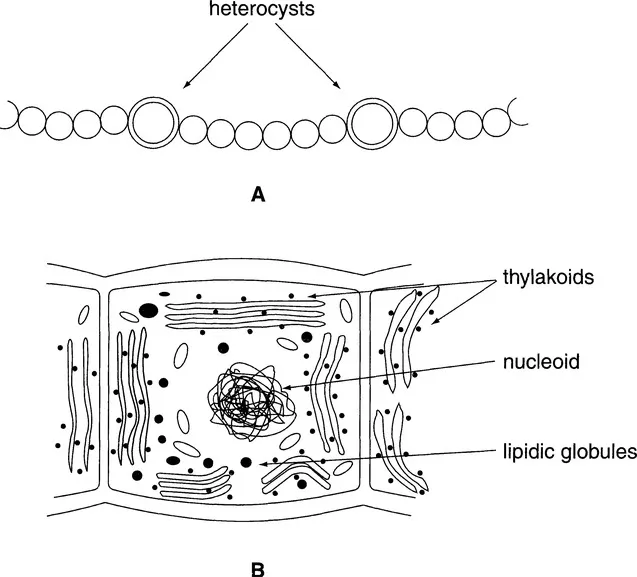
Some bacteria are also photosynthetic because of the presence of chlorophyllian pigments (bacteriochlorophylls). At this frontier of the plant kingdom are found the Cyanobacteria or blue bacteria, which were earlier called blue algae. They are bacteria because they have no nuclear compartment but are classified among the algae on the basis of the vegetative organization, which can be as evolved as that of other algal groups (nematothallus and cladothallus). These cyanobacteria are also distinguished by the differentiation of a complex photosynthetic structure, the chromoplasm, which associates concentric membranes and pigments dissolved in lipidic drops (Fig. 1.1). They are perfectly autotrophic for carbon. The genera Nostoc and Oscillatoria are representative of these organisms.
Fig. 1.1. (A) Colony of a cyanobacterium, nostoc, with its heterocysts. (B) Details of a cell with its chromatoplasm composed of thylakoids, nucleoid, and lipidic globules.
With the animal kingdom
Although there is no difficulty in recognizing the kingdom to which a rabbit or a carrot belongs, the limits are fluid and some groups can pose problems. Three examples are taken up below.
The Euglena are generally considered unicellular green algae. They are autotrophic because of the presence of numerous plasts enclosing pigments associated with membranes. However, they are mobile, because they do not have the usual cell wall found in plant cells and do have a flagellar part. When cultured in conditions highly favourable to their multiplication, the population grows exponentially but the rate of nuclear division is higher than that of chloroplast division. Thus, there is a gradual diminution in the number of plasts per cell. This phenomenon recurs until the Euglena has only one chloroplast at the time of mitosis. The result is an unequal division: one of the cells inherits a chloroplast and the other does not and becomes the source of an aplastidiate cell line. If this line has a source of carbon in its culture medium, it multiplies as do protozoa in culture and thus belongs irreversibly to the animal kingdom.
Some organisms at the frontier of the two kingdoms are similar to one another, but some are photosynthetic and some are not. This is the case of many flagellates as well as some algae, the Pyrrophycophytes, and protozoa belonging to the Rhizopods. The two groups have the same type of alternation of generations. An alternation of generation, although typical of plants, characterizes the cycle of trypanosome, a protozoa that causes malaria.
Another problem is that of the Mycophytes or fungi, which play a very important role in biotechnology. Even though they are not chlorophyllian, they are generally considered very similar to plants because of their fixed life, their alternation of haploid-diploid phases, the formation of meiospores, their cell wall, or their plant-like organization, which puts them in the category of Thallophytes. However, their heterotrophy, the presence of chitin in the composition of their cell wall, the existence of centriolar parts to direct mitosis, and a mobile plasmode phase in the group Myxomycetes (also called Mycetozoa by zoologists) are arguments that can be opposed to botanists who insist on considering those organisms as plants. It is preferable, in fact, to consider the fungi an independent and original group between plants and animals.
d) The major plant groups and biotechnology
It is presently difficult to propose a classification of plants that would be accepted by the entire community of botanists. There is a constant evolution, not to mention a constant revolution, in the science of systematics. Therefore, though our position may be obsolete, it has the merit of being still accepted by most people and compatible with most studies.
In the system followed in this book, plants are divided into two major groups depending on whether they possess an axis that provides support, distribution of water and metabolites, and an erect habit. This axis (or trunk) allows us to distinguish the Cormophytes or higher plants from Thallophytes or lower plants. The following dichotomies are used to make subdivisions within these two groups.
within the Thallophytes:
- Algae,
- Fungi,
- Lichens.
within the Cormophytes:
- Cryptogams:
* Bryophytes or Mosses and related plants,
* Pteridophytes or Ferns and related plants,
- Prephanerogams: Ginkgoales and Cycadales,
- Phanerogams:
* Gymnosperms or Conifers and related plants
* Chlamydosperms: Gnetales and related plants,
* Angiosperms:
Monocotyledons or Liliopsida,
Dicotyledons or Magnolopsida.
Only some of these groups or classes of plants are of interest in biotechnology. We will have occasion to note this as we describe the technical approaches and bioindustrial uses of these plants. We can, however, as a preliminary approach, characterize the relationships between these various groups and biotechnology and report the following general indications.
The Algae or Phycophytes now constitute a field of application of biotechnological research involving experiments to obtain cultured organisms, cells and calluses the commercialization of some raw materials designed for animal and human food, additives (agar, alginates, carrageenans), and products that have pharmaceutical (particularly antiviral) and cosmetic applications. Genetic engineering is still in its infancy in this group.
The Fungi or Mycophytes, particularly yeasts and Ascomycetes, are of particular interest in genetic engineering and biotechnology. The entire genome of bakers’ yeast (Saccharomyces cerevisiae) has been sequenced and most of it has been decoded. We will see that these yeasts are most valuable auxiliaries in acquiring knowledge about and mastering plant genomes.
In the Cormophytes, the biotechnological approach is still very weak with respect to the Bryophytes (mosses and related plants) and Pteridophytes (ferns and related plants). In the Prephanerogams, the Ginkgo presently holds a great deal of interest because of the presence of certain diterpenes with significant pharmacological effects. For similar reasons, much attention has been given to Gymnosperms (conifers and related plants) and especially Taxus (yew), which yields taxol, an extract with known antimitotic and thus antitumoral properties. The synthesis of such products by genetic engineering is important in current research. It is probable that the equally active search for pharmacodynamic molecules will lead the pharmaceutical industry to look at Chlamydosperms and Ephedra in particular.
The Phanerogams are by far the most interesting group in terms of biotechnological approaches and, consequently, the most often cited group in this book. It includes almost all plants of agronomic and horticultural interest and is, of all living organisms after bacteria and yeasts, the group in which genetic engineering seems to have advanced most and for which economic considerations seem to have the greatest weight.
1.1.2. Plant growth and development
a) Ontogenic programmes
The structure of a plant corresponds to a set of morphological and physiological phenomena that occur in a precise order and exploit information inscribed in the genetic inheritance of each individual: this is called an ontogenic programme. In fact, there are two successive programmes in plants, since there is an alternation of generations: a sporophytic ontogenic programme for the plant strictly speaking and a gametophytic ontogenic programme during the appearance of reproductive processes. Each programme comprises processes of growth and development.
Growth is characterized essentially by a phase of intense cell multiplication followed by a phase of cell elongation. The first is meresis and the second is auxesis. It is essentially an increase in biological mass by true reproduction of cells.
Development corresponds to the successive appearance and differentiation of the organs of the plant: stems, branches, roots, leaves, and flowers. The differentiation of organs requires highly varied modes of cell and tissue differentiation.
The progress of an ontogenic programme requires an initial element or structure that can vary greatly in plants. This element is very often unicellular: a zygote for the sporophyte and a spore for the gametophyte. It can also be multicellular: fragments of tissues or organs as well as similar parent cells of gametes that could give rise directly, by parthenogenesis and true multiplication, to a new organism, without participating in a phenomenon linked to sexual reproduction.
Usually, there are two periods in the life of a plant: growth and development of vegetative parts and then acquisition of the capacity to reproduce sexually, which is the building of a reproductive apparatus. Like any living organism, the plant then ages and dies and its matter returns to the mineral world through processes of humification before being cycled again by the development of new organisms.
b) Growth and development of vegetative parts
The growth and development of vegetative parts generally begin with the fertilization and formation of a zygote, the modalities of which will be discussed later. Let us note, however, that there is an apparent paradox here, since there is normally an opposition between the concepts of ...
Table of contents
Cover
Half Title
Title Page
Copyright Page
Table of Contents
Preface
Introduction
1 Some Basic Concepts of Biology
2 Plant Biotechnology and Genetic Engineering
3 Applications of Biotechnology and Genetic Engineering
4 Plant Biotechnologies and Bioethics
Conclusion. The Future of Biotechnology and Genetic Engineering in Plants
Exercises
Glossary
Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Genetic Engineering and Biotechnology by Yves Tourte in PDF and/or ePUB format, as well as other popular books in Ciencias biológicas & Biología. We have over 1.5 million books available in our catalogue for you to explore.