The movement of organisms is a fundamentally important ecological process. Voluntary movement is a critical aspect of animal biology and ecology. Humans have been keenly interested in the movement of individual animals and populations for millennia. Over 2000 years ago, Aristotle wrote about the motion of animals, and the associated philosophical and mathematical concepts, in his book, De Motu Animal-ium (Nussbaum 1978). Historically, it was critical to understand how and where wild food sources could be obtained. Thus, early humans were natural animal movement modelers. In modern times, we are interested in the movement of animals for scientific reasons and for making decisions regarding the management and conservation of natural resources (Cagnacci et al. 2010).
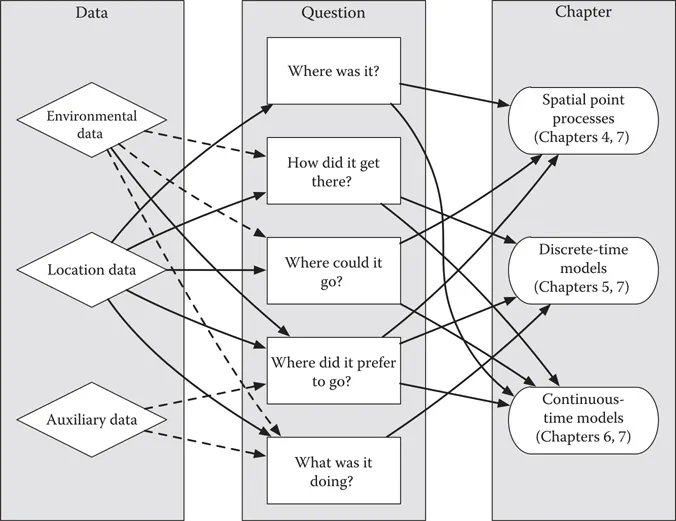
The study of wild animals can be challenging. Animals are often elusive and reside in remote or challenging terrain. Many animals have learned to minimize exposure to perceived threats, which, unfortunately for us, include the well-intentioned biologist approaching them with binoculars or a capture net. Therefore, it is no surprise that the development of animal-borne telemetry devices has revolutionized our ability to study animals in the wild (Cagnacci et al. 2010; Kays et al. 2015). Animal telemetry has helped us overcome many of the practical, logistical, and financial challenges of direct field observation. Telemetry data have opened windows that allow us to address some of the most fundamental ecological hypotheses about space use (“Where is the animal?”), movement (“How did the animal get there?,” “Where could it go?”), resource selection (“Where does the animal like to be?”), and behavior (“What is the animal doing?”) (Figure 1.1).
1.1 BACKGROUND ON ANIMAL MOVEMENT
Animal movement plays important roles in the fitness and evolution of species (e.g., Nathan et al. 2008), the structuring of populations and communities (e.g., Turchin 1998), ecosystem function (Lundberg and Moberg 2003), and responses to environmental change (e.g., Thomas et al. 2004; Trakhtenbrot et al. 2005; Jønsson et al. 2016). The scientific study of animal movement has a deep history, and we are unable to explore all of the ecological implications and methodological developments in a single volume. Instead, we focus on several specific inferential methods that can provide valuable ecological insights about animal movement and behavior from telemetry data.
The importance of animal movement in larger-scale ecosystem function probably inspired the Craighead brothers to develop and deploy the first radio collars on grizzly bears (Ursus arctos) from Yellowstone National Park in the 1960s (Craig-head and Craighead 1972). Satellite tracking devices are now capable of pinpointing animal locations at any moment, remote sensing provides ever refined environmental data, and biotelemetry tags allow for the simultaneous collection of important physiological and behavioral information from wild animals. These technological advances will lead to a better understanding of how individual decisions affect demographic parameters and ultimately translate into population dynamics. In this sense, animal movement can provide the long-sought bridge between behavior, landscape ecology, and population dynamics (Lima and Zollner 1996; Wiens 1997; Morales et al. 2010; Kays et al. 2015).
In what follows, we provide a brief summary of research findings, existing knowledge, and analytic approaches for important aspects of animal movement ecology. We organized these topics into 10 sections:
Population dynamics
Spatial redistribution
Home ranges, territories, and groups
Group movement and dynamics
Informed dispersal and prospecting
Memory
Individual condition
Energy balance
Food provision
Encounter rates and patterns
1.1.1 POPULATION DYNAMICS
In classical models of population dynamics, predators and prey encounter each other in proportion to their overall abundance over space and reproductive rates decrease as the global population density increases. This is because traditional models of population and community dynamics assume we are dealing with many individuals that are well mixed (Turchin 2003). Such “mean field” representations of population dynamics can provide good approximations when the physical environment is relatively homogeneous and organisms are highly mobile, or when organisms interact over large distances. However, when the external environment or the limited mobility of organisms results in lack of mixing, the conditions experienced by a particular member of a population or community can be quite different from the mean (Lloyd 1967; Ovaskainen et al. 2014; Matthiopoulos et al. 2015). That is, when per capita vital rates are affected by varying local conditions, the observed population and community dynamics can differ markedly from mean field predictions.
Population dynamics involve births, deaths, immigration, and emigration; modern tracking technology, together with new statistical models, can greatly improve our understanding of these processes. The individuals that comprise a population can vary in several traits and individual behavior can change in response to internal and external stimuli. Individual traits and behavior determine the way they interact with the environment and other organisms while the conditions that individuals experience ultimately translate to their performance (i.e., growth, survival, and reproduction).
Survival analysis can be used to model changes in hazard with time and in relation to covariates such as location, age, body condition, and habitat type. Detailed tracking through satellite telemetry enables spatial information and survival data to be combined at small temporal scales, leading to an increasingly sophisticated understanding of the determinants of survival (Murray 2006; Haydon et al. 2008; Schick et al. 2013). Likewise, changes in movement behavior can be used to infer reproductive events in some species (Long et al. 2009). However, to take full advantage of these data, new analytic techniques should take into account the sequential nature of individual survival and reproduction. For example, the chance of an animal dying of starvation depends on its history of encounters with food items and foraging decisions.
Coupling demographic data with movement models is an area of active research, but is still somewhat nascent. Spatial capture–recapture (SCR) models provide a way to formally connect animal encounter data with movement processes; we refer the interested reader to Royle et al. (2013) and references therein for additional details. The methods presented in this book will be critical for formally integrating location data and demographic data in future SCR modeling efforts.
1.1.2 SPATIAL REDISTRIBUTION
Classical reaction–diffusion models, such as those used by Fisher (1937) to describe the spread of an advantageous mutation within a population assume that mortality and recruitment rates depend linearly on local population density and that individuals move at random over a large and homogeneous area. Early implementations of these models were also used to describe the dynamics of population invasion and range expansion (e.g., Skellam 1951; Andow et al. 1990; Shigesada and Kawasaki 1997), and later, were embedded in a hierarchical statistical modeling framework (e.g., Wikle 2003; Hooten and Wikle 2008; Hooten et al. 2013a) to provide inference about spreading populations.
Diffusion equations have been justified as a good approximation to the displacement of individuals performing a “random walk.”* Although we know that animals do not move at random, the diffusion approximation can still be sufficient at certain (usually large) scales and also serves as a null model to compare with more complex models (Turchin 1998).
More general forms of movement can be taken into account by formulating spatial population models as integral equations. These have commonly been formulated in discrete time, yielding integro-difference equations where local population growth is combined with a “redistribution kernel” that describes the probability that an individual moves from its current location to another one in a given time-step.† The temporal scale of these models is usually set to match reproductive events so that the redistribution kernel represents successful dispersal rather than regular movement. A great deal of theoretical and empirical work has explored the consequences of kernel shape, particularly in the tail of the distribution, on invasion speed (Kot et al. 1996; Powell and Zimmermann 2004).
There are many ways to make spatial population models more realistic and appropriate for particular species, places, and scales of interest. A good starting point is to consider the spatial structure of the population, which is generally accepted as an important prerequisite for more accurate ecological predictions (Durrett and Levin 1994; Hanski and Gaggiotti 2004).‡ The spatial structure of populations can range from classical closed populations to a set of s...