
- 408 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Ecology and Palaeoecology of Benthic Foraminifera
About this book
This is an important and authoritative review of foraminiferal ecology, the first for over a decade. Professor Murray relates ecological data on living forms of foraminifera to the palaeoecology of fossil species, and defines in detail areas of global distribution.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
EcologyIndex
Biological SciencesChapter 1
Introduction
Ecology is the study of the causes of patterns of distribution and abundance of organisms. It is concerned with interactions between individuals and their physical and chemical environment, interactions between species and also interactions between individuals of the same species. The approach to ecology may be through field studies, laboratory experiments or mathematical modelling. Ideally ecological studies involve all three approaches because each provides new data or a new outlook which give rise to further questions or new ideas. During the 1970s the thrust of marine ecology moved away from the exploration of new habitats towards understanding more of the interrelations between organisms within the benthic community and with the overlying waters (Holme and McIntyre 1984), e.g. the ecosystem approach.
Distribution studies of benthic foraminifera have been carried out for well over a century, but a truly ecological approach did not develop until 1952 when Walton introduced the rose Bengal staining method for differentiating between protoplasm-containing (i.e. living or recently living) and empty (i.e. dead) tests. Now field studies of benthic foraminiferal ecology consider not only the living individuals but also the postmortem taphonomic processes which influence the preserved dead assemblages. This is because the principal application of ecological data on modern foraminifera is to aid the palaeoecological interpretation of fossil assemblages.
In spite of the large quantity of literature on modern benthic foraminifera there is still much work to be done to establish the basic distribution patterns. Most studies are field based; only a few involve laboratory experiments or mathematical modelling. However, the introduction of new techniques, such as the stable isotope composition of the test wall, and ultrastructural and biochemical studies of the soft parts, is throw ing new light on their physiology and mode of life.
Although some foraminifera live on hard substrates such as shells or rocks, or attached to plants or other organic structures, most live on or in soft sedimentary substrates. In theory, competition between species for space and food may be ecological controls, but in practice there are no data to suggest that either takes place among small benthic sediment inhabitants. Most infaunal communities can undergo large increases in density without evidence of competitive exclusion Dayton, in Strong et al. 1984).
The niche
Lee (1974: 209-10) suggests that:
The concept of niche is not easily understood except in very abstract terms. Often micropaleontologists and protozoologists think of niche in a spatial or habitat sense and commonly only from data gathered at a single point in time. While these studies are valuable in themselves, they give only very little insight into the biotic constraints which are important in the dynamic processes which establish the realized niches of foraminifera.
Further (Lee 1974: 219)
… we really know very little about the realized niches of any group of foraminifera from any locality in the world. Hopefully in the future we will have enough field and laboratory data to describe the realised niches of a few foraminiferal species. It staggers the imagination to consider just the field work involved….
Factors influencing the occurrence of organisms may be abiotic (temperature, salinity, dissolved oxygen availability, pH, etc.) or biotic (food, intra- and inter-specific competition, predation, etc.). Muller (in Lee 1974) attempted to quantify some of these parameters for four species of foraminifera and he developed a polygonal graphical plot to represent the niche. The six most important factors are plotted along vectors, the centre being displaced from the centre of the circle in order to emphasize the importance of the three abiotic factors. He inferred that Ammonia beccarii occupies a broader niche (44%) than Spiroloculina hyalina (27%) (Fig. 1.1). The latter has a low interspecific competition coefficient and occurs in low numbers in the field. Ammonia beccarri is widely distributed and this is consistent with its niche breadth.
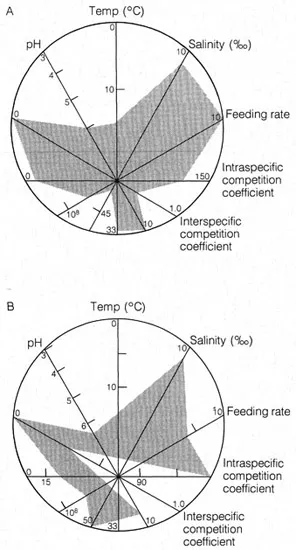
Figure 1.1 Graphic representations of the realised niches of (A) Ammonia beccarir, (B) Spiroloculina hyalinea (based on Lee 1974)
The niche concept is most applicable to micro-habits. It still remains a theoretical ideal rather than a practical objective, indeed, it may prove more useful to know broad distribution patterns rather than the minutiae of the niche of each species, for the interaction of environmental controls may cause the realised niche to vary from one region to another.
Environmental variability
It is likely that abiotic environmental parameters are of greater importance than biotic factors in shaping the communities which live in variable environments such as marginal marine settings. They also play a major role in delimiting biogeographic divisions.
Because many benthic foraminifera approach the size of detrital material in sedimentary substrates, they are very difficult to study experimentally under natural conditions. There is no evidence of strong interspecific competition for a potentially limiting resource at meiofaunal level (Warwick 1981; Dayton, in Strong et al. 1984). However, parasites (including viruses, bacteria and fungi) are considered to play a major role in shaping communities, even on a biogeographic scale (May, in Strong et al. 1984).
Environmental disturbance may be an important control on meiofaunal organisms. Apart from bottom disturbance from the action of waves or currents there may be localised disturbance by organisms. Fish, and especially rays, dig pits in the substrate to seek prey. In so doing, they displace the microfauna some of which may be eaten by ‘picker’ fishes. Apart from that, the disturbed patches may need to become recolonised by the microfauna so that the sea floor becomes a mosaic of successful patches (Dayton, in Strong et al. 1984). This process is not easy to quantify and it introduces variability in some environments which may otherwise seem to be stable. Similarly, the presence of tubes or burrows of macrofaunal species may provide niches for small species such as foraminifera. On ribbed bivalve shells, the grooves rather than the ribs are the parts colonised by clinging or fixed species (Plate 3c).
Diversity and abundance
Diversity refers to the number of species, but to be meaningful a diversity index must also take into account the number of individuals per species. One view is that greatest diversity is achieved in those assemblages in which all the species are equally abundant. In nature such a pattern is rarely achieved. In assemblages with few species one is normally more abundant that the others and this can be readily shown by histogram plots of the rank order of species (Fig. 1.2A, B). As the number of species increases the percentage abundance of the dominant species decreases (Fig. 1.2C, D). In these examples diversity is expressed by two diversity indices: the Fisher a index and the information function H(S) (see Appendix A for a description of them).
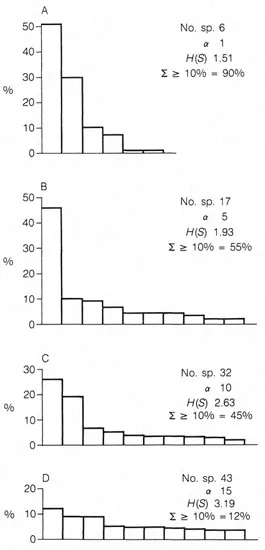
Figure 1.2 Histogram plots in rank order of the most abundant 10 species in assemblages of differing diversity. (A) Living, sta. 8, Long Island Sound (Buzas 1969); (B) living, sta. 1253; (C) dead, sta. 1245; (D) dead, sta. 1248, North Carolina Shelf (Murray 1969)
Synthesis of data
A principal objective of this book has been to synthesise the large volume of published data so that patterns may be defined which will assist the palaeoecological interpretation of fossil assemblage.
Most papers concern a relatively small area of sea floor and often the author has sought differences within the area rather than emphasising similarities. For example, Medioli et al. (1986) studied a small area of the Nova Scotia continental shelf and recognised seven assemblages based on factor analysis, but Williamson et al. (1984) who studied a larger area had a single (Eggerella advena) assemblage in place of the seven. In this book similarities are stressed.
The starting-point for ecological studies is the distribution of living assemblages for they can be related to environmental parameters. On death, or as a consequence of reproduction, empty tests are added to the sediment to give the dead assemblage. Consideration of the processes which influence the composition of the dead assemblage is important for understanding the preservation of ecological signals in the fossil record. Detailed discussion of this is given in Chapter 5.
Ecological studies are carried out at species level but much useful information can be gained by grouping data. Although fossil assemblages commonly consist of species which are no longer extant, genera are longer ranging through geological time. Some of the general attributes of modern assemblages are applicable to fossil examples. The methods of data analysis are discussed in Appendix A and include general attributes such as diversity, measured by the two commonly used methods of the information function H(S) and the Fisher a index, the ratio of the three wall structure types (agglutinated, porcellaneous, hyaline), and generic predominance.
In considering the distribution of benthic foraminifera by geographic area it is most meaningful to recognise basic recurrent associations which, for convenience, are named according to the one or two key species. Such associations are most objectively defined by multivariate analysis and indeed this is possible within limited areas. However, it is not feasible to take the entire data set for a whole continental margin and to subject it to multivariate analysis.
The associations recognised here are based on the dominant species in terms of percentage abundance in either living, dead or total assemblages. The common additional species are those present in >10% abundance in some or all of the samples. It is a matter of observation that the common additional species differ from one geographic area to another and this reflects varying patterns of biogeographic distribution.
The data used are only from samples in which >100 individuals were counted as this is considered to be the minimum acceptable size. Because of this restriction the interpretations reached here sometimes differ from those of the original authors of the data. Not all papers give essential ecological data, but an attempt has been made to summarise basic environmental information for each of the associations.
The recognition of associations based on the dominant species might be taken to imply that each is discrete and has a sharp boundary with the next. This is clearly not the case for, as already noted, the common additional taxa vary from one region to another so the concept embraces variability. Also, on a local scale different associations may show seasonal replacement. Even with the often crude generalisation of environmental parameters, it is normally the case that the abiotic field differs from one association to another (e.g. see Fig. 6.4).
Objectives and structure of the book
Probably 99% of the papers on the ecology and distribution of modern benthic foraminifera are written by geologists. This is primarily because of the palaeoecological utility of foraminifera. In this book the objectives are to provide a state-of-the-art synthesis of the enormous quantity of available data, to provide it in a form which make it readily applicable to understanding the fossil record, and to demonstrate the major biogeographic patterns and their ecological causes.
Chapters 2-5 deal with the important aspects of life processes, stable isotope studies, population dynamics and the relationship between living and dead assemblages. These set the scene for a consideration of the geographical and ecological distribution of species and associations (Chs 6-7). These results are summarised in Chapter 18. As far as possible factual data have been concentrated into tables and figures and into the four appendices.
The section on palaeoecology is short because it is considered necessary only to define basic principles and to discuss some case-st...
Table of contents
- Cover
- Half Title
- Title
- Copyright
- Contents
- Preface
- Acknowledgements
- Chapter 1 Introduction
- Chapter 2 Life processes
- Chapter 3 Stable isotope studies
- Chapter 4 Population dynamics
- Chapter 5 Relationship between living and dead assemblages
- Chapter 6 Atlantic seaboard of North America
- Chapter 7 Gulf of Mexico and Caribbean Sea
- Chapter 8 Atlantic seaboard of South America
- Chapter 9 Atlantic seaboard of Europe and Africa
- Chapter 10 Atlantic Ocean – summary and comparison
- Chapter 11 The Mediterranean
- Chapter 12 Indian Ocean
- Chapter 13 Western margin of the Pacific Ocean
- Chapter 14 Eastern margin of the Pacific Ocean
- Chapter 15 Pacific Ocean
- Chapter 16 Southern Ocean
- Chapter 17 Arctic Ocean
- Chapter 18 Summary of modern distribution patterns and characteristics of assemblages
- Chapter 19 Palaeoecology
- Appendix A Methods
- Appendix B Ecological data for selected genera
- Appendix C Faunal reference list – recent taxa
- Appendix D Glossary
- References
- General index
- Geographic index
- Taxonomic index – suprageneric categories
- Taxonomic index – genera and species
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Ecology and Palaeoecology of Benthic Foraminifera by John W. Murray in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Ecology. We have over 1.5 million books available in our catalogue for you to explore.