ICPA provides a forum for researchers and academics who share a common interest in ecological psychology to come together, present new research, and foster ideas towards the advancement of the field. This volume is the fourteenth in the Studies in Perception and Action book series, and highlights research presented at the 19th International Conference of Perception and Action (ICPA) in the summer of 2017. Since 1991, this edited book series has appeared in conjunction with the biennial ICPA meeting. The short papers and empirical articles presented in this book represent the contributions of researchers and laboratories from across the globe. The reader will find new, cutting-edge research on a wide variety of topics in perception and action. This volume will especially appeal to those that are interested in James J. Gibson's ecological approach to psychology, as well as, more broadly, students and researchers of visual and haptic perception, perceptual development, human movement dynamics, human factors, and social processes.

eBook - ePub
Studies in Perception and Action XIV
Nineteenth International Conference on Perception and Action
- 124 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Studies in Perception and Action XIV
Nineteenth International Conference on Perception and Action
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Subtopic
Cognitive Psychology & CognitionIndex
PsychologyChapter 1:
Action and Coordination
Different Muscle Synergies Required for the Reconstruction of Muscle Activation Patterns During Point-to-Point Movements With and Without a Rod
Raoul M. Bongers, Tim A. Valk, & Leonora J. Mouton
University of Groningen, University Medical Center Groningen, Center for Human Movement Sciences, Groningen, The Netherlands
University of Groningen, University Medical Center Groningen, Center for Human Movement Sciences, Groningen, The Netherlands
Although the degrees of freedom problem has been clearly articulated by Bernstein (1967) almost a century ago, the views on how this problem is solved to perform goal-directed actions differ to a large extent. Central in all these accounts is how to organize the redundant degrees of freedom (DoFs) in such a way that the system becomes controllable, that is, how to get rid of the control of all individual DoFs. Synergy is a notion commonly used for this. A primary challenge for all these accounts is that synergies are a stable coordination among DoFs, yet allow for flexible task performance. For instance, many different reaching movements can be used to reach the same target.
One account on synergies prominent in the literature claims that muscles are orchestrated in fixed, but functional groups, called muscle synergies (d’Avella, Saltiel, & Bizzi, 2003). The idea then is that muscles that are active at the same time during a movement, e.g., because they perform a similar action around a joint, are united in muscle synergies. Within a muscle synergy the proportional activation of each muscle is fixed. Muscle synergies can then be used as building blocks of movements, to produce muscle activation patterns in all muscles that they incorporate. Activating muscle synergies in a time-varying way and combining the activation of different synergies allows to produce a wide variety of movement with a given set of muscle synergies. However, if this is the case, actions performed are constrained by a fixed set of muscle synergies, then this fixed and limited set of muscle synergies should be able to generate the activity of muscles to perform one task in different ways. That is, different types of reaching movements should have to be explained with the activation of a restricted set of muscle synergies.
To examine the explanatory value of this account, in the current study we employed tool use to assess the extent to which a fixed set of synergies is used to achieve the same task in different ways. To produce the same end-effector trajectory as in reaching without a tool, alternative movements and thus alternative muscle activation patterns are required to bring the tip of the end-effector on the target. We examined whether these alternative muscle activation patterns for the different tools can be reconstructed from the same, fixed set of muscle synergies.
Method
Fifteen right-handed participants made point-to-point movements from a central position to one of eight peripheral targets, displayed on a table in front of the participants. The targets were equally spaced on a circle around the central position with a radius of 25 cm. Participants made both center-out movements, from the central position to one of the peripheral targets, and out-center movements, from one of the peripheral targets back to the central position. These movements were made with different end-effector length, i.e., with the index finger, and with rods of 5 cm, 15 cm, and 25 cm attached to the index finger.
In total, each participant performed five blocks of 64 trials each, resulting from sixteen different movement directions, i.e., the center-out and out-center movements towards and from eight targets, and four different end-effector lengths. Both the order of movement direction and used end-effector within a block, as the order of blocks, were presented in a randomized order.
During these movements, the muscle activation patterns of 22 muscles in the lower arm, upper arm, trunk and back were recorded at 1600 Hz. Raw data were band-pass filtered (4th order Butterworth filter, 20-450 Hz), rectified, and low-pass filtered (4th order Butterworth filter, 10 Hz) to determine the linear envelope of the EMG signal. For these linear envelopes, the part from 200 ms before movement onset until 200 ms after movement termination was selected, normalized in time and amplitude, and averaged over trials, and taken for further analyses.
Subsequently, per end-effector length, the sixteen processed signals, one for each movement direction, were used to determine a set of muscle synergies for every end-effector length. This was done using a nonnegative matrix factorization algorithm, in which the error between the modelled muscle activations, as produced by a set of synergies W and their time-varying coefficients C, and the observed muscle activations, is minimized.
To test whether the muscle synergies extracted from every end-effector length could produce the same muscle activation patterns, the extracted muscle synergies from one condition were used to reconstruct the observed muscle activation patterns from another condition. Using a nonnegative reconstruction method this was done for all pairs of conditions. In this case, the time-varying coefficients C were determined with respect to a fixed muscle synergy set W from another end-effector condition. The quality of reconstruction was determined by computing the explained variance of this reconstruction.
Results and Discussion
The data of one participant has been fully analyzed and is presented here. Preliminary data analyses of the other participants suggest similar trends.
On average, the synergies extracted from every end-effector condition explained 79.2% of data variance of the observed muscle activation patterns. For this participant in all conditions the analyses revealed four synergies. As can be seen in Figure 1, the observed muscle activation patterns from every condition were reconstructed well by muscle synergy combinations from that condition. In this figure can also be seen that different activations of muscles were used for the different end-effector lengths.
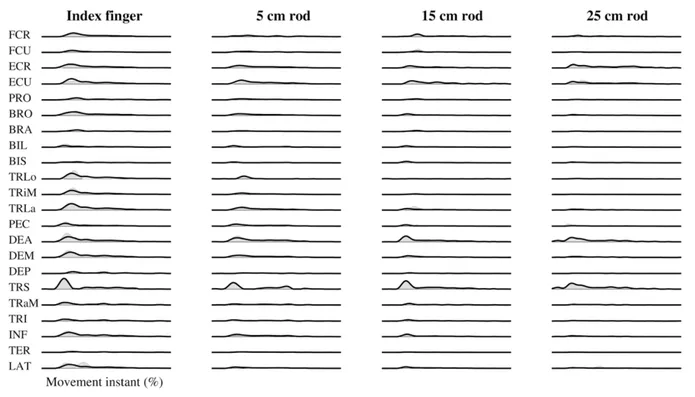
Figure 1. Example of reconstruction of muscle activation patterns during a pointing movement in a forward direction. Black lines represent the reconstruction by the combination of muscle synergies, whereas the grey areas represent the observed muscle activation patterns.
The main question of this study was whether these different muscle activation patterns could be produced by the same set of muscle synergies. Results showed that the muscle activation patterns from a certain end-effector condition were reconstructed best with the muscle synergies from that specific end-effector condition. Importantly, the explained variance dropped substantially when muscle synergies of another condition are used (Figure 2). Moreover, the explained variance of the reconstruction became worse as the difference between the lengths of the end-effector between conditions became longer.
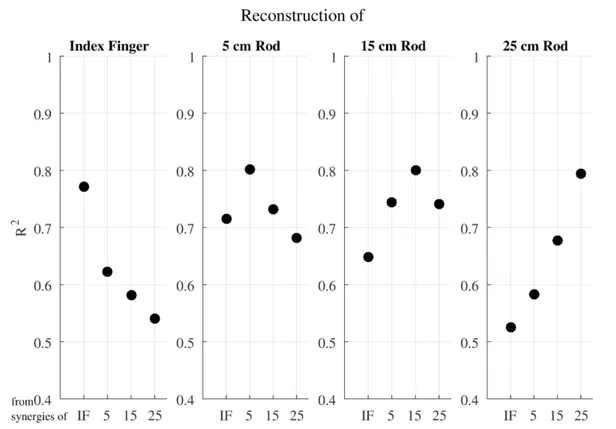
Figure 2. Explained variance for the reconstruction of the EMG data from one condition using the muscle synergies from the same or another condition.
The analyses so far indicated that a different set of muscle synergies is required to reconstruct muscle activation patterns for different conditions. Hence, our preliminary findings do not seem to support the idea that a fixed set of muscle synergies are used as building blocks to construct a wide variety of movements. Therefore, the current findings suggest to us that the muscle synergy approach is not capable to explain the flexibility of muscle recruitment in reaching tasks with a tool. We suggest that approaches more able to explain flexibility in synergies, such as notions based on a dynamic systems account (Turvey, 2007) or the UCM approach (Latash, Scholz, & Schöner, 2007), should be taken as a starting point to understand synergistic organization of DOFs.
References
Bernstein, N. A. (1967). The coordination and regulation of movements. Oxford: Pergamon Press.
d’Avella, A., Saltiel, P., & Bizzi, E. (2003). Combinations of muscle synergies in the construction of a natural motor behavior. Nature Neuroscience, 6(3), 300–308.
Latash, M. L., Scholz, J. P., & Schöner, G. (2007). Toward a new theory of motor synergies. Motor Control, 11(3), 276–308.
Turvey, M. T. (2007). Action and perception at the level of synergies. Human Movement Science, 26(4), 657–697.
Hysteresis Effects When Grasping an Object With a Simple Hand Prosthesis
Raoul M. Bongers1, A. Heerschop1, & C.K. van der Sluis2
Persons with a traumatic injury and/or amputation of the upper limb are often fitted a prosthesis to replace the missing limb and to restore action capabilities. Technological advancements in the last two decades have definitely improved the capabilities of the hands, which have improved from simple grasping devices to multi-articulated hands that can perform different hand postures and grip types. Despite these technological developments the rate of use of these prostheses has been relatively low. Therefore, training of the user has to be taken to the next level to enable the user to take full advantage of the possibilities of the prosthetic device. Most studies take as a starting point the idea that the neuromotor system must (a) form an internal model of the prosthetic hand and (b) to learn to produce the requisite control signals. The current study is the first to exploit whether a dynamical systems account of prosthesis use is viable.
Different types of active prostheses are on the market. The current study focuses on myoelectric prostheses. In this type of prosthesis electric motors control motions around the joints such as the wrist, as well as the movements of the digits. These motors are controlled through myo-signals produced by muscles in unaffected parts of the body or residual musculature in the residuum. Importantly, for the majority of the prosthetic users the multi-articulated prosthesis is controlled with two myo-signals, usually picked up from the flexors and extensors of the wrist. If multiple grip modes are to be controlled with two signals, switching between the grip modes is required. Once within a grip mode, the grip/hand can be closed and opened by ...
Table of contents
- Cover
- Title
- Copyright
- Contents
- Preface
- Meeting History
- Scientific Committee
- Contributors
- Chapter 1: Action & Coordination
- Chapter 2: Interpersonal Coordination
- Chapter 3: Perception and Perceptual-Motor Learning
- Chapter 4: Perception of Affordances
- Chapter 5: Postural Control and Coordination
- Author Index
- Keyword Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Studies in Perception and Action XIV by Julie A. Weast-Knapp,Gert-Jan Pepping in PDF and/or ePUB format, as well as other popular books in Psychology & Cognitive Psychology & Cognition. We have over 1.5 million books available in our catalogue for you to explore.