
- 664 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Plant Toxicology
About this book
In order to keep track of all the compounds and pathogens affecting plant metabolism and development, you would need to spend all your waking hours combing periodicals and the Internet in dozens of languages, as new toxins via pollutants and migratory or mutant pathogens are being discovered every day.
Plant Toxicology, Fourth Edition start
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Toxicology1 Characteristics of Plant Life: Hazards from Pollutants
Bertold Hock and Nicola M. Wolf
Technische Universität München, Freising, Germany
I. GREEN PLANTS AS PHOTOAUTOTROPHIC ORGANISMS
Living organisms are characterized by their extraordinary complexity. It is manifested in the higher plant by a hierarchy of structures comprising organs, tissues, and cells. The following chapters provide an introduction to plant organization starting with the cell as the smallest elementary unit. Emphasis is laid upon plant-specific features, which are discussed with respect to their susceptibility to environmental contaminants.
II. FUNCTIONAL ORGANIZATION OF THE CELL
A. Role of Membranes
Plant cells as eukaryotes are characterized by their compartmentation into membrane-enclosed reaction spaces. This structure separates different metabolic pathways but at the same time allows a grouping of connected biochemical functions. This process allows sophisticated regulations.
The borders of compartments are composed of biomembranes (Fig. 1). These thin and highly flexible structures determine the architecture of biological systems. Biomembranes are flat, asymmetrical structures, which are closed and usually topologically equivalent to the surface of a sphere or a torus. Along with the basic composition of lipids and proteins there are variety of individual compositions. The ratio of proteins to lipids varies within a range of 1:4 to 4:1. Both components are held together by hydrophobic interactions. The lipids form a bimolecular layer, which serves as a barrier and prevents the passage of polar molecules. Integral proteins penetrate completely or at least partly the lipid bilayer. Conspicuous examples are the chlorophyll a/b-binding protein of chloroplast membranes and the adenosine triphosphate (ATP) synthase of mitochondrial and chloroplast membranes. Peripheral proteins bind at the membrane surface to integral proteins from which they can be detached easily by appropriate solvents. They include clathrin, spectrin, and ankyrin of the plasma membrane. Usually membrane proteins have mediating functions: they serve as receptors, ion channels, ATP-powered pumps, or transporters. In many cases enzymatic activities are crucial for the function. This explains why the two sides of the membrane bilayer are usually different.
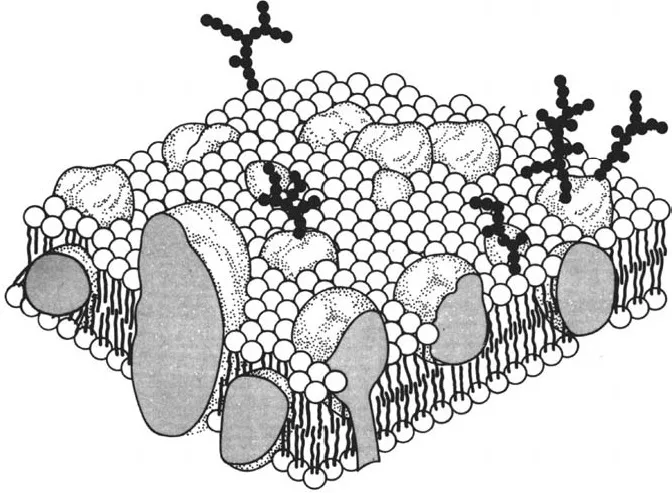
Figure 1 Biomembrane with integral and peripheral membrane proteins (1). The protruding oligosaccharide chains belong to glycoproteins and glycolipids.
Transport through a biomembrane uses one of the three following mechanisms (Fig. 2):
1. Passive diffusion: Only a few substances are able to penetrate the lipid bilayer. Examples are gases such as oxygen (O2), nitrogen (N2), or carbon dioxide (CO2) and some small, uncharged molecules such as ethanol, urea, or benzene, which dissolve easily in the lipid bilayer. In principle this also holds true for water (H2O) although its diffusion may be accelerated by transport proteins (aquaporins). Passive diffusion of H2O through the biomembrane is crucial for osmosis. In contrast, lipid bilayers are practically impermeable to charged molecules.

Figure 2 Scheme of the membrane transport (2). Passive transport follows an electrochemical gradient as free diffusion or facilitated diffusion and is mediated by transport proteins. In contrast, active transport requires adenosine triphosphate– (ATP-)powered pumps and moves ions or small molecules uphill against the electrochemical gradient.
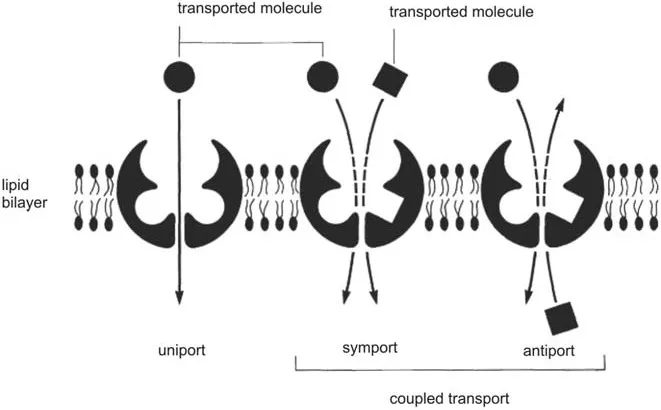
Figure 3 Scheme of transport proteins, which act as uniporters, symporters, or antiporters (2).
2. Facilitated diffusion (catalyzed diffusion): Similar to passive diffusion this mechanism does not require energy and leads only to a concentration equilibrium. However, transport proteins (channels and transporters) are required for this type of membrane transport. Three options are available (Fig. 3): (a) unidirectional transport (uniport), which moves only one kind of molecule; (b) symport (cotransport), in which two molecules or ions are transported in the same direction, and one of them, for instance, H+, follows a concentration gradient; (c) antiport, in which two molecules or ions move in opposite directions, one of them following a concentration gradient. Both antiporters and symporters mediate coupled reactions in which the energetically unfavorable reaction is coupled to an energetically favorable reaction.
3. Active transport: The process pumps ions or small molecules through a membrane against a chemical concentration gradient and/or electric potential. This ATP-powered transport moves ions such as H+, K+, Ca2+, and Na+ in one direction. It is mediated by adenosine triphosphatases(ATPases). The required energy is released by the hydrolysis of ATP to adenosine disphosphate (ADP) and inorganic phosphate (Pi). This active transport indirectly drives symport and antiport.
The compartmentation of cells creates huge membrane areas, which offer extended targets to environmental compounds and toxins approaching and penetrating the cells. Binding of a variety of substances can lead not only to impairment of individual functions, but also to destruction of whole compartments. The abolition of compartmentation always leads to cell death.
Compartmentation entails a separation of protoplasmic and nonprotoplasmic spaces: the plasmalemma separates the protoplast from the cell wall at the outside, the tonoplast from the vacuole at the inside (Fig. 4). The plasmalemma (plasma membrane) allows the movement of solutes into the protoplast as well as outward. In addition, it is involved in signal transduction between the outside and inside and mediates hormonal as well as light effects. This allows the cell to detect changes of its environment and react accordingly. For this purpose several receptor types as well as redox chains are available. A prominent component of the plasmalemma is the H+-ATPase. This transport ATPase plays a crucial role in the uptake of nutrients and pH regulation. By means of this ATP-driven H+ pumping the membrane becomes energized, and the electrochemical potential for the import of solutes through ion channels and carriers is maintained by ATP hydrolysis. This function is taken in animal cells by the sodium pump (Na+/ K+-ATPase). Whereas in this case the pump is usually coupled to K+ import and Na+ export, proton coupling is used by plants. The proton pump transports a single H+ for each hydrolized ATP and creates in this way a large electrical potential up to -300mV (inside the membrane negative) as well as a proton gradient. The values reach a pH of c. 7.1 at the inner side of the membrane and at the outside values between 4.5 and 5. A comparison of the different strategies is illustrated by Fig. 5.
Cells with intensive active transport such as root hairs require between 25% and 50% of their total cellular ATP to keep their proton pumps running. In addition hydrogen adenosine triphosphatases (H+-ATPases) play an important role in cell elongation (cf. acid growth theory: Chapter 1, 2D structure and function of the cell wall).
The plasma membrane contains a multitude of further membrane proteins, depending on the specific cell. In addition to H+-ATPases, cation and anion channels as well as carriers for sucrose, nitrate, and amino and a multitude of hormone and blue light receptors have been identified. Even many pollutants act on components of the plasmalemma, especially on H+- ATPases. Many xenobiotics also enter the cell, where they are taken up directly or after conjugation to glutathione into the vacuole. In other cases cytochrome P450–dependent hydroxylations occur, followed by further metabolic steps.
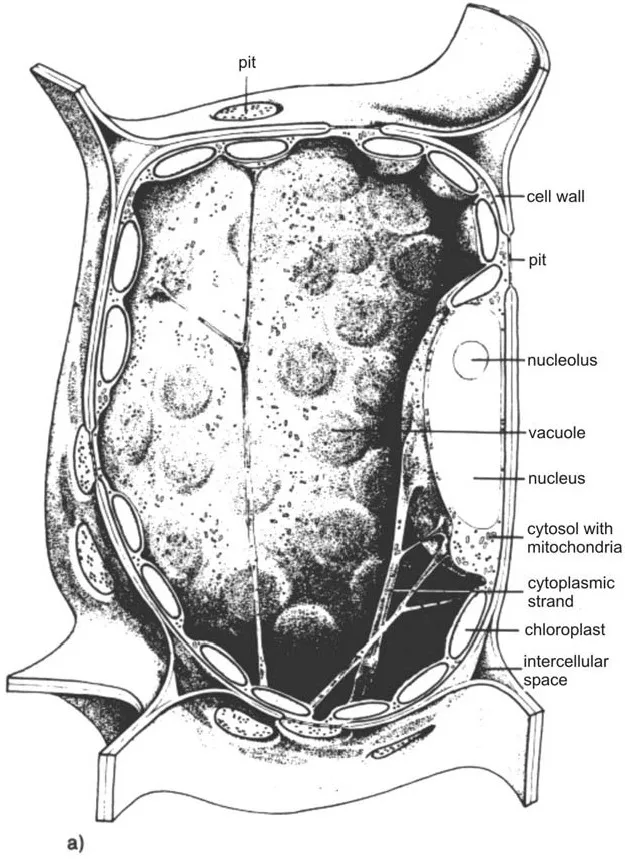
Figure 4 Structure of a plant cell: (a) Scheme (3); (b) electron micrograph of a tobacco mesophyll cell. ER, endoplasmic reticulum. (With kind permission of Prof. Katherine Esau, University of California.)
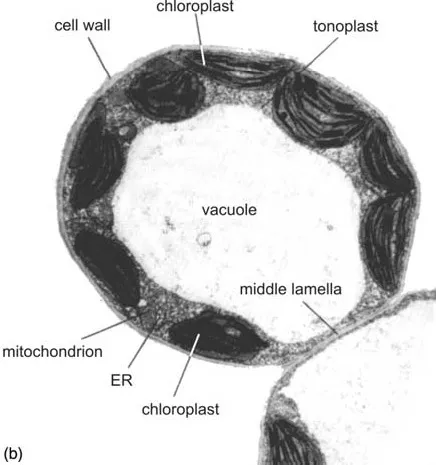
Figure 4 Continued.
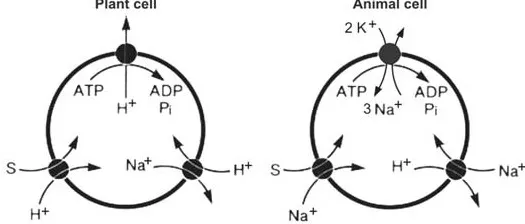
Figure 5 Different strategies of plant and animal cells regarding the uptake of solute (S) and pH regulation. ATP, adenosine triphosphate; ADP, adenosine diphosphate; Pi, inorganic phosphate. (From Ref. 4.)
According to the principle of hierarchic structuring the protoplast is divided into several compartments. The membrane-enclosed spaces are organelles in a narrower sense. There are organelles bounded by two membranes (cell nucleus, mitochondria, and plastids) and organelles surrounded by a single-membrane envelope (such as endoplasmic reticulum, dicytosomes, lysosomes, peroxisomes, glyoxysomes, and microbodies). These organelles are embedded into the cytosol, which also contains additional particles (organelles in a wider sense such as ribosomes as well as fibrous elements belonging to the cytoskeleton). Sometimes the traditional grouping of the protoplast into cell nucleus and cytoplasm is used.
With respect to cell compartmentation the following rules are important: an envelope composed of two membranes separates plasmatic from other plasmatic spaces (P spaces). The P spaces are characterized by their potential to synthesize nucleic acids and/or proteins. The delimitation of the nucleus, mitochondria, and plastids against the cytosol falls into this group. A delimitation by a single membrane separates plasmatic from endoplasmatic spaces (E spaces). The latter spaces are not capable of nucleic acid or protein synthesis. The cell wall and vacuole are nonplasmatic spaces, as are the lumen of the endoplasmic reticulum (ER), dicytosomes, lysosomes, and microbodies. The symbiont theory of organelle evolution explains these distinctions: mitochondria and plastids are derived from prokaryotic cells that have colonized as endosymbionts, the progenitors of modern eukaryotes.
Cell membranes are subjected to remarkable dynamics, to which membrane flow substantially contributes. Membrane flow is mediated by an intensive vesicle stream within the cytosol. There are several options, depending on the respective cell: the pathway from the ER, which is connected to the nuclear envelope, via the Golgi apparatus to the plasmalemma (exocytosis) or to the lysosomal compartment. On the other hand, components of the plasmalemma, e.g., receptors, are recycled by endocytosis. By means of membrane flow large amounts of newly synthesized fatty acids and lipids are transported f...
Table of contents
- COVER PAGE
- TITLE PAGE
- COPYRIGHT PAGE
- PREFACE
- CONTRIBUTORS
- 1. CHARACTERISTICS OF PLANT LIFE: HAZARDS FROM POLLUTANTS
- 2. PLANT STRESS: AVOIDANCE, ADAPTATION, DEFENSE
- 3. UPTAKE AND TRANSPORT OF XENOBIOTICS
- 4. AIR POLLUTION: TRACE GASES AS INDUCERS OF PLANT DAMAGE
- 5. LIMITATION OF SALT STRESS TO PLANT GROWTH
- 6. MINERAL ELEMENT TOXICITIES: ALUMINUM AND MANGANESE
- 7. HERBICIDES
- 8. MOLECULAR BASIS OF TOXIC EFFECTS: INHIBITION OF CELLULAR PATHWAYS AND STRUCTURAL COMPONENTS
- 9. METABOLISM AND ELIMINATION OF TOXICANTS
- 10. HOST–PATHOGEN RELATIONS: DISEASES CAUSED BY VIRUSES, SUBVIRAL ORGANISMS, AND PHYTOPLASMAS
- 11. INTERACTIONS BETWEEN HOST PLANTS AND FUNGAL AND BACTERIAL PATHOGENS
- 12. ALLELOPATHY
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Plant Toxicology by Bertold Hock, Erich F. Elstner, Bertold Hock,Erich F. Elstner in PDF and/or ePUB format, as well as other popular books in Medicine & Toxicology. We have over 1.5 million books available in our catalogue for you to explore.