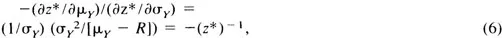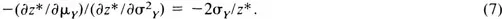The sixth volume in this respected series systematically presents and evaluates quantitative models of various foraging phenomena, including: steady state decision rules; acquisition of decision rules; perception and learning in foraging behavior.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
The foraging economy of most animals includes elements of stochastic variation. Learning may reduce the uncertainty associated with problems of information, but the uncertainty induced by risk may be independent of the forager’s experience. In the first case the forager may be uncertain about resource quality, so that the acquisition of information becomes important. By sampling available alternatives, the forager can update its probabilistic assessments of profitability (Green, 1980; Kacelnik, 1979; McNamara, 1982; Oaten, 1977). For this reason, an animal’s foraging efficiency may depend on its sampling efficiency (Houston, Kacelnik, & McNamara, 1982; Krebs, Kacelnik, & Taylor, 1978; Lima, 1984). Several contributions to this volume indicate the scope and intensity of current interest in ecological questions concerning information in foraging processes.
In stationary environments, (i.e., when resource quality is not a function of time), or when a forager can recognize distinct resource types (e.g., different prey species) prior to exploitation, sampling is less important. In effect, the organism may acquire “total information.” However, the problem of risk remains when the outcome of a foraging decision is a random variable; that is, the forager may possess full information concerning the probability distributions of benefits and costs associated with available options, but the variation in those benefits and costs may still impose survival (and sometimes fecundity) risk (Caraco, 1980; McNamara & Houston, 1982; Pulliam & Millikan, 1982; Real, 1981; Stephens & Charnov, 1982). This chapter deals solely with this second aspect of foraging in stochastic environments.
To study preference over levels of risk, we initially characterize a probability distribution of a foraging currency by its statistical moments: mean, variance (or standard deviation), and skew. We usually take reward size as the random variable in experiments. But the logic we develop applies to problems where either the cost of obtaining a required amount of food (Caraco, 1981a) or the rate of energy intake (Stephens & Charnov, 1982) is the random variable of interest.
For each member of a set of benefit or cost distributions, we relate the statistical moments to the probability that the forager will obtain less than its physiologically required intake in the time available for feeding. We take the probability of an energetic deficit as the proxy attribute for fitness in our models. For nonbreeding animals this probability is functionally related to the probability of starvation (Caraco, Martindale, & Whittam, 1980; Pulliam & Millikan, 1982). We assume that natural selection favors discrimination abilities (e.g., Commons, 1981; Commons, Woodford, & Ducheny, 1982) and decision rules that promote survival in nonbreeders. Therefore, a forager should always prefer the benefit distribution associated with the smallest attainable probability of an energetic deficit (Caraco et al., 1980; Houston & McNamara, 1982; Pulliam & Millikan, 1982; Stephens & Charnov, 1982). Furthermore, these probabilities should allow one to predict a preference ranking over available foraging options (Caraco, 1983). Perhaps the most interesting departure of the theory of risk-sensitive foraging from deterministic foraging theory is the relationship predicted between choice and energy budgets. The risk-sensitive forager’s preference for one reward versus another need not be fixed but can depend (in the manner described in the following) on a comparison of required and expected food intake (Caraco et al., 1980; Houston & McNamara, 1982; Stephens, 1981; for a related phenomenon see de Villiers & Herrnstein, 1976).
A Minimization Premise
The models presented here employ optimization, though it is in no way anticipated that optimal behavior will be commonly realized. Optimization provides a logical, quantitative means to generate testable hypotheses based on assumptions about natural selection. Properly interpreted, optimization models make no unreasonable expectations of nature. The purpose of such models is to elucidate the consequences of possible selective forces governing Darwinian evolution (Hinson & Staddon, 1983; Maynard Smith, 1978; Pyke, Pulliam, & Charnov, 1977).
Suppose an animal exploiting a stochastic environment has n foraging opportunities during a finite time interval (a day for convenience). Denote the reward at trial i (i = 1, 2, …, n) with the independent random variable xi. The mean and variance of each xi, are finite: E[xi], V[xi] < ∞.
is the total reward acquired during the day. Then
, and
. F(Y) is the distribution function; F(y) = Pr[Y ≤ y]. As long as one xi does not dominate the sum, and the xi’s are not uniformly skewed (see following), Y should approach normality for sufficiently large n by the central-limit theorem.
Assume that the forager must accumulate a total reward exceeding R to satisfy daily physiological requirements. Let F(R) = Pr [Y ≤ R] be the probability of an energetic deficit. For simplicity we assume that selection on survival might favor minimizing this probability. More complex relationships between energy intake and fitness (Caraco, 1980; McNamara & Houston, 1982) are discussed later.
Following Stephens and Charnov (1982), we use a simple transformation as a useful characterization of the forager’s problem. Because Y approaches normality, the random variable z, where z = (Y − μY)/σY, is approximated by the standard normal distribution. Minimizing F(R) is then equivalent to minimizing
Suppose the forager (fully informed by assumption) must choose to allocate its time to one of k elements (that is, reward probability distributions) of a set S1, S1, might consist of k foraging habitats situated so that the cost of switching habitats during the day would be prohibitive. In making the choice, larger means are always attractive, because ∂ϕ(zR)/∂μY < 0. The influence of the standard deviation depends on the sign of (R − μY); that is,
When a forager can expect its intake to exceed its requirement (μY > R, a positive energy budget), increasing variance decreases an option’s value. But when the forager can expect an energetic deficit (μY < R, a negative energy budget), increasing reward variance enhances the value of an option (Caraco et al., 1980; Houston & McNamara, 1982; Pulliam & Millikan, 1982; Stephens & Charnov, 1982).
We can immediately form some hypotheses from (2) and (3). Consider an animal presented with experimental choices between a constant reward (σY = 0) and a variable option (σY > 0) with an expected value equal to the constant reward. According to (2), the animal should prefer the constant reward if its expected energy budget is positive. However, according to (3), the animal should prefer the variable option if its expected energy budget is negative. These predicted behaviors usually are termed, respectively, risk-aversion and risk-proneness (Keeney & Raiffa, 1976), because risk ordinarily is assumed to depend on a measure of variability (e.g., Pollatsek & Tversky, 1970).
Next, presume an animal is presented with a series of choices constructed from all combinations of the elements taken two at a time from a set S2; that is, if S2 contains k elements, one considers the k!/2[k− 2)!] different choice situations. Each reward probability distribution fi
S2 has the same expected value (and no skew) and a unique standard deviation. Because the assumed overall objective is the minimization of ϕ (zR), i.e.,
expressions (2) and (3) predict that the animal’s preference ranking over the elements of S2 should exhibit a strictly monotone ordering (Caraco, 1983). For positive energy budgets (μY > R), preference rank should decrease strictly as σY increases about the common mean. If the forager can expect a negative energy budget (μY < R), preference rank should increase strictly as the standard deviation increases. In either case, the minimal ϕ(zR) should be preferred over all other outcomes, but the preference ordering reverses if the sign of the quantity (R − (μY) is changed.
The analysis of the preceding paragraph yields the same predictions if risk is characterized in terms of reward variance. However, the minimization of ϕ(zR) and variance-discounting models (see following) make different predictions in terms of the interaction of mean reward with variability. Suppose the forager can choose across a large number of options that differ in both mean reward and standard deviation (under the assumption of no skew). The distribution(s) preferred over all others should be the (μY, σY) combination(s) minimizing the probability of an energetic deficit; that is, the distributions should be ranked from minimal to maximal ϕ(ZR) for a particular energy budget. For fixed R, consider a set of reward distributions yielding the same Pr[Y ≤ R]. The forager should be indifferent between (i.e., prefer equally) any two of these distributions, because they enhance survival equally (which assumes that indifferences will be transitive). Because the F(R) are equal, each such distribution must have the same z value (say z*) according to (1). Then the various (μY, σY) elements of this indifference set determine the indifference curve (Stephens & Charnov, 1982):
Given a preference ordering in terms of ϕ(zR), an indifference set defines a straight line in the μ−σ plane. The marginal rate of substitution of σY for μY along this indifference curve is
a constant. Linearity of indifference curves holds whether the forager avoids or prefers reward variability. The sign of the slope of (5) is negative in the first case and positive in the second, and z* has the sign opposite to the sign of the slope of the indifference curve.
If indifference curves are linear in the μ–σ plane, mean and variance
must trade off nonlinearly. Equation (5) implies that the forager that ranks options in terms of ϕ(zR) should exhibit decreasing risk-sensitivity (Keeney & Raiffa, 1976). Analyzing the marginal rate of substitution of variance for mean along an indifference curve (constant z*) shows that
Therefore, the trade-off between μY and
depends on the value of the mean, because σY fixes μY for a given R and z*. The consequences of (7) are the following:
For a positive energy budget (μY > R), a forager should accept a large...
Table of contents
Cover
Half Title
Title Page
Copyright
Contents
About the Editors
List of Contributors
Preface
Introduction
Part I: Aspects of Optimal-Foraging Theory
Part II: Foraging in a Changing Environment
Part III: Operant and Pavlovian Processes
Part IV: Special Foraging Adaptations
Part V: Methodology
Author Index
Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Foraging by Michael L. Commons, Alejandro Kacelnik, Sara J. Shettleworth, Michael L. Commons,Alejandro Kacelnik,Sara J. Shettleworth in PDF and/or ePUB format, as well as other popular books in Psychology & Experimental Psychology. We have over 1.5 million books available in our catalogue for you to explore.