The usual method for studying mental processes entails taking words in linguistics -- or concepts in logic -- and establishing the connections and relationships between them. Thus, the traditional approach to semantic problems -- those of meaning and understanding -- is through language. Most researchers agree that thought and language are generated by deep-seated semantic structures determined by the structure of the brain. Until now, however, all attempts at constructing semantic models have been made on the basis of linguistic material alone, without taking brain structure into account. Analysis of these models shows them to be as inadequate as those based on the method of the black box.
This book approaches the problem of the organization of higher psychological functions a different way -- by analyzing the functional organization of the neural structures that gradually form universal categories from "raw" sensory material. At the higher levels of the brain's operation, these universals correspond to the basic categories of thought and language. The visual system provides rewarding material for such an approach, both because it is relatively well researched and because it is the main source of sensory information in humans. With this in mind, this monograph examines the whole process of the transformation and description -- the coding of visual information. The most important aspect of this process is the transition from the description of visual space to the description of individual objects and the relationships between them. This transition is made possible by the existence in the visual system of various mechanisms that developed during evolution as a result of environmental influences.
Written for a wide circle of investigators in disciplines associated with different aspects of the functioning of the brain -- physiologists and psychologists -- this book is also of importance to engineers and mathematicians working on the problems of artificial intelligence, and linguists and philosophers interested in the deep structures that form the universals of thought and language.

- 290 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
1
Processing of Visual Signals in the Retina and the LGN
The overview of information processing in the retina and LGN in this chapter does not pretend to be complete and is confined only to those aspects that are essential for solving the problem, as formulated in the Introduction. This includes the operations the retina and LGN must perform to make manageable the main jobs of the visual brain: pattern recognition and the description of the spatial parameters, both within an image and between images. Therefore, not every step in the account in this chapter is accompanied by formal references. The assertions made here are based on classic papers and reviews by Hartline (1940, 1959), Kuffler (1952), Kuffler and Nicholls (1976), Rodieck (1965), Dowling and Werblin (1969), Robson (1975), Barlow and Mollon (1982), Trifonov (1982), and Shapley and Enroth-Cugell (1984).
The retina and LGN form the visual subcortex through which visual information is transmitted to the cortex. The immediate source of the visual input to the cortex is the cells of the LGN, where the description of the image—characterized as the distribution of excitation among cells—corresponds in some way to the distribution of luminance on the retina delivered by the optic nerve. This process is not a simple transmission from point to point, however. The image is essentially transformed, and knowledge of the operations that accomplish this transformation is necessary for an understanding of the higher functions of the visual cortex.
The information processing at this level involves the description of the image by the multiplicity of receptive fields (RFs) in the subcortical areas. The neurons of the retina and the LGN measure the integrated light energy in the central summation zone of their receptive fields. The existence of an inhibitory periphery causes the neurons at this level to carry out a series of preliminary processing operations: separating the signal from surrounding noise, emphasizing contours and high spatial frequencies, and producing spatial and temporal decorrelations of the image. However, the description of the visual field at this level is still made on a retinotopic, or point-to-point, principle: The distribution of excitation in the on- and off-neurons of the retina and in the neurons of the LGN corresponds to the distribution of luminance in the receptors or to the distribution of positive and negative contrasts relative to the average luminance level. Because of the appearance of separate on- and off-systems, a foundation is created for the transition to local spectral analysis at the next level, in the striate cortex.
The first transformation of the image delivered by the optical system of the eye on the receptors of the retina is performed by the triad, photoreceptor—horizontal cells—bipolar cells, in which there is active feedback within and between layers (Fig. 1.1a). This triad provides an example of cells interacting with each other by means of graded depolarizations and hyperpolarizations, rather than through all-or-none impulses arriving at synaptic terminals. Increasing the amount of light that falls on receptors causes an increase in the hyperpolarization of the cell. In the process of transmission from the receptor cell to the bipolar cell, the signal is transformed by horizontal cells, acting as interneurons, which mediate lateral interaction. The RF of the horizontal cell is large: 0.3–1.0 min of arc, depending on the species. Horizontal cells sum excitation from a great number of photoreceptors. In addition, the horizontal cells become synchronized, so that excitation of even one cell can influence others. Therefore, by summing the excitations, the horizontal cells can exert their influence as they make contact between receptor cells and bipolar cells. Although all receptors respond to light by hyperpolarizing, the same does not hold true for bipolar cells. Some bipolars hyperpolarize when illuminated, whereas others depolarize.
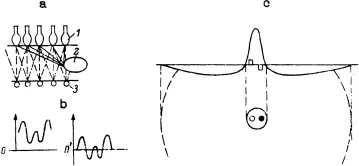
FIG. 1.1. Scheme of retinal input: a, scheme of triad: photoreceptor (1), horizontal cells (2), bipolar cells (3); b, displacement of zero level of signal; c, scheme of weighting function (above) and RF (below) of bipolar cells. Light and dark stimuli are shown by columns on the weighting function, and by circles on RF shown in the diagram.
Thus, an on-and-off process begins at the level of the bipolars. The end arborizations of the terminals of bipolars are localized in the inner plexiform layer (Fig. 1.2), which consists of two sublayers, a and b. Sublayer a contains the endings of off-bipolars, which respond by hyper-polarization to a spot of light; sublayer b contains the endings of depolarizing on-bipolars. Thus, dendritic stratification is essential to the organization of an on-and-off process. Dendritic trees of on-ganglion cells (localized in lamina b of the inner plexiform layer, and of off-cells located in lamina a) contact the terminals of bipolars.
Amacrines and interplexiform cells are interneurons that modulate the influence of bipolar cells on ganglion cells. The role of these interneurons (horizontal cells and amacrines) is to create the spatial organization of the RF. The RFs of cells of visual subcortex through which the information flows (bipolars, ganglion cells, and cells of the LGN) have an incomplete, but essentially round, center-surround form (Spinelli, 1966).
In on-center cells, an increase in light elicits an excitatory response in the center (i.e., an increase in firing rate in the ganglion cells or geniculate cells and depolarization in bipolars) and an inhibitory response in the surround. In off-center cells, a decrease in light elicits an excitatory response in the center and an inhibitory response in the surround.
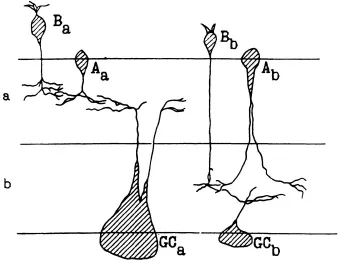
FIG. 1.2. The arrangement of cells in the inner plexiform layer: a and b, sublayers: A, amacrines; B, bipolars; GC, ganglion cells (from Famigletti, Kaneko, & Tachibana, 1977). Copyright © 1977 by the AAAS. Reprinted with permission.
According to the model that is generally accepted, this organization may be explained in terms of an overlapping center–surround mechanism. Each mechanism can be conceived of as containing the cells of the preceding level and the interneurons; their signals are pooled and converge with different signs onto the cell at the next stage in neural processing. The profiles of activity of both the excitatory and the inhibitory mechanisms are of Gaussian form. The diameter of the inhibitory mechanism is 4 times greater than that of the excitatory one.
Because of the algebraic summation of the spatially organized excitatory and inhibitory processes, the resulting profile has the form of difference of Gaussian (DOG). The profile of activity, or the weighting function (WF) in a unidimensional direction, is defined as:
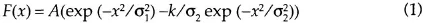
What operations are performed by such a retinal construction? One of the operations the retina performs is retinal adaptation. The purpose of adaptation is to keep the visual response the same when the level of illumination changes; in other words, adaptation allows invariant description, despite changes in illumination. This invariance, as discussed earlier, is a central problem in pattern recognition.
Mach (1865) was the first to understand this problem. He contended that the ratio of the quantity of light in the background and on the image remains constant under different conditions of illumination. Thus, we can state that contrast is independent of illumination. Contrast, by definition, is measured in physical terms and depends only on the reflectances of object and background; thus, it is independent of level of illumination. We now discuss how the calculation of contrast can be explained in terms of retinal adaptation.
The eyes of the human function in a wide range of luminances—about 11 orders of magnitude. Any further increase in contrast does not change response, due to saturation. Retinal adaptation allows us to use a limited dynamic range of visual cells for the description of signals in all the ranges of illumination.
The second operation performed by the retina is the formation of separate on-center and off-center systems. On-center and off-center systems are separated in the retina and in the LGN, and the signals from them are united only in the cells of the visual cortex (Norton & Sherk, 1984). The existence of on-center and off-center systems is a necessary condition for the linear operations in the visual cortex; they are the main mechanisms underlying pattern recognition. To clarify, let us discuss the role and organization of linear mechanisms in the transmission of information in the visual system.
Linear systems are the simplest mechanisms that allow for the coding of information in a form that permits full restoration. This does not mean, of course, that the image on the retina is restored in the visual cortex. It means only that the information is transmitted without loss.
Linearity means that the response to a stimulus, which may be represented as the sum of several stimuli, is equal to the sum of the responses to these stimuli recorded separately. The most complete way to represent the linear properties of the cell is by determining its weighting function. For simplicity, let us consider the unidimensional case. If x defines the position of a point on the retina, and F(x) is the function of the stimulus characterizing the value of luminance in x, the response of the linear cell is:

where the interval a–b is the position of the RF on the retina, and Ω(x) is the weighting function characterizing the cell. It is obvious that if R1 and R2 are responses to stimuli F1 and F2, then the response to stimulus F(x) = F1(x) + F2(x) would be R = R1 + R2.
As stated earlier, the existence of two systems is a necessary condition of linearity in visual cortex. The reason is as follows: In a linear process, the response to two light spots should be equal to the algebraic sum of the responses to each spot. For example, if the center of a round RF (Fig. 1.1c) is stimulated simultaneously by light and dark spots, the response is equal to zero. In an on-center RF, one spot produces the excitation, while the other one produces inhibition. This is the situation in a bipolar RF, where graded hyperpolarizations and depolarizations are summed on the membrane. In cells where the input to the synapse is an impulse, however, it is impossible to receive both excitation and inhibition from the same cell of the preceding stage. In this case, the inhibition on the membrane is produced with the aid of the interneuron. I show in the next chapter that linearity in regions of the RF of linear cortical cells is produced by on- and off-geniculate cells whose RFs coincide spatially, such that one cell provokes excitation and the other one provokes inhibition via the interneuron. Therefore, light and dark spots shown in the subfield of the RF do not evoke a response.
Enroth-Cugell and Robson (1966) showed that the type of ganglion cells called X display linear spatial summation. When they are tested with sinusoidal gratings, a position can be found where a null response occurs. The center of the ganglion cell is linear, because it is formed by bipolars. However, there are doubts as to whether the inhibitory surround also has linear properties. The inhibitory Gaussian distribution of ganglion cells is organized by amacrines. These cells, in contrast with bipolar cells, sometimes transmit signals by impulses, rather than only by graded potentials. In such cases, the excitatory part of the membrane potential, its depolarization, is reflected in the response of the amacrines; inhibition is not transmitted. Because the amacrine is the interneuron that inhibits the transmission of the signal to the ganglion cell, it follows that the surround zones cannot be excited by a stimulus opposite in sign to the stimulus that excites the center.
At first glance, this assertion is paradoxical. It is well known, for example, that the periphery of an off-center ganglion cell may be excited by turning on a light in the peripheral circle or by turning off such a light in an on-center cell. In other words, the surround behaves as a linear part of the RF. We can explain this fact as follows: Application of the light to the surround evokes a change in the mean level in the RF (Fig. 1.3b). In such a case, the luminance in the center of the RF is decreased relative to the mean level; that is, the center is stimulated by a dark spot. It is not clear if we should designate this as the effect of the RF surround on the gain of the center. There is considerable evidence that the area of adaptation in the RF is either equal to or smaller than the signal summation area of the RF center itself (for a review, see Shapley & Enroth-Cugell, 1984). However, many authors have proposed that the center’s gain ought to be regulated by the surround. In their review, Shapley and Enroth-Cugell found that only X-cells were studied in the photopic range, and there was a small but significant increase of center gain when the adapting spot was enlarged to cover the surround. It has been established that the latency of the response in the surround is greater by about 100–200 msec than the latency of the response at the center to a spot. This supports the treatment of the experiment in Fig. 1.3 as adaptation, because the response occurred at th...
Table of contents
- Cover
- Halftitle
- Title
- Copyright
- Contents
- Introduction
- 1 Processing of Visual Signals in the Retina and the LGN
- 2 The Receptive Fields of the Striate Cortex
- 3 The Modules of the Visual Cortex: Neurophysiological Investigations
- 4 Piecewise Fourier Analysis: Psychophysical and Behavioral Evidence
- 5 The Primary Segmentation of Visual Space
- 6 The Prestriate and Peristriate Cortices and Their Functions
- 7 The Two Basic Visual Mechanisms
- 8 Pattern Recognition and the Functions of the ITC
- 9 The Description of Spatial Relations and the Functions of the PPC
- 10 Hemispheric Asymmetry and Visual Recognition
- 11 A Hypothesis and Model of the Higher Divisions of the Visual Brain: The Double Dichotomy of the Visual Brain
- 12 A Perceptual Model of the World
- Epilogue
- References
- Author Index
- Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Vision and Mind by Vadim D. Glezer in PDF and/or ePUB format, as well as other popular books in Psychology & Cognitive Neuroscience & Neuropsychology. We have over 1.5 million books available in our catalogue for you to explore.