
eBook - ePub
The Neuroscience of Handwriting
Applications for Forensic Document Examination
- 267 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Neuroscience of Handwriting
Applications for Forensic Document Examination
About this book
The Daubert trilogy of U.S. Supreme Court cases has established that scientific expert testimony must be based on science grounded in empirical research. As such, greater scrutiny is being placed on questioned document examination generally, and handwriting comparison in particular. Bridging the gap between theory and practice, The Neuroscience of
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.

I
Fundamental Aspects of Motor Control and Handwriting

1
Neuroanatomical and Neurochemical Bases of Motor Control
Introduction
The human brain is a complex system governing automatic and willed behaviors, multimodal perception, emotion, and restorative functions. It is without doubt that brain function today is the refinement of millions of years of adaptation and evolution. With few exceptions, the nervous system control functions we observe in humans today can be traced to corollary functions of lower animals. For example, there is considerable evidence demonstrating a relationship between cranial capacity and hand morphology and function over the past 1.75 million years. Fine motor control of the hand and articulatory system for speech are perhaps the most obvious among the many evolutionary advances that can be traced to an increase in brain size and complexity.
Historical Perspective on Brain Function for Hand Motor Control
Numerous writings can be found in the literature on brain function throughout antiquity. Much of this literature is nicely summarized in a very readable treatise by Stanley Finger (1994). The idea that different parts of the brain subserved different functions may be traced to the writings of the Roman physician Galen (AD 130–200). Galen’s anatomical work with various animals showed that the cerebrum was softer than the cerebellum, leading to his conclusion that motor and sensory pathways were separate. He further reasoned that, unlike the motor nerves, sensory nerves needed to be pliable to retain the sensory information for long periods of time. Galen thus asserted that the sensory nerves went to the cerebrum while the motor nerves went to the cerebellum because the former was softer than the latter.
Ventricular localizationalists dominated brain science throughout the fourth and fifth centuries. Figure 1.1 depicts the neuroanatomical understanding of brain localization of the 1200s as envisioned by Albertus Magnus (1206–1280). Throughout the years following Galen, the dominant theory held that higher brain functions such as cognition, imagination, and memory were associated with the cerebral ventricles. The ventricles were where the spirits from the sensory nerves ended before being taken up by the motor nerves to invoke action.

Figure 1.1 Drawing of the ventricles by Albertus Magnus published in the 1506 edition of Philosophia naturalis. (Photo source: Corbis, with permission.)
Functional localization in the ventricles was widely accepted for hundreds of years, even into the Middle Ages. For example, in 1481, the Italian physician Antonio Guainerio described two patients: one who was unable to speak more than a few words at a time (a condition we now refer to as aphasia) and another who could not remember people’s names. Assuming both conditions stemmed from a memory disorder, Guainerio diagnosed their problems as stemming from excessive buildup of phlegm in the posterior ventricle. It was not until the early 1500s that the ventricular doctrine began to unravel. During this time, Leonardo da Vinci dissected hundreds of brains from cadavers and conducted experiments on ventricles from cattle brains. His observations were largely inconsistent with the assertions held by the ventricular localizationists of the time. While reasoning that the flow of “nervous spirit” from sensory nerves should be more midline than lateral, daVinci fell short of openly challenging the doctrine that higher mental functions were seated in the ventricles.
Others, however, argued for a completely different view of the cerebral functional localization and the role of the ventricles. Andreas Vesalius (1514–1564) rekindled interest in brain function during the Renaissance. At age 23, Vesalius received a grant (in the form of material) from the Senate of the Republic of Venice to conduct public dissections. Books on his work published in 1543 set the stage for a dramatic paradigm shift in the structure–function relationship of the human brain. Vesalius’s main argument was that since the human ventricular system was not different in shape from other mammals and since other animals lacked higher reasoning powers, how could these powers be relegated to the ventricles? In describing Galen’s work, he uncovered nearly 200 cases in which Galen’s anatomic drawings were incorrect. While generally opposed to the idea of ventricular localization, Vesalius did not reject the traditional view that animal spirits were produced in the ventricles. His progressive stance on anatomy was dissociated from his adherence to traditional principles of physiology. More than 1,300 years after Galen, Vesalius wrote that the ventricles are no more than spaces into which air flows to be mixed with vital spirit from the heart and then transformed into animal spirit distributed through the nerves to organs of sensation and motion.
A century after Vesalius’s death, Thomas Willis (1621–1675) published a book entitled Cerebri Anatome in which he proposed that the cerebral gyri controlled higher cognitive functions. Vital and involuntary functions were attributed to the cerebellum (along with what we now refer to as the midbrain and pons). The corpus striatum was thought to play a role in sensation and movement. Willis had effectively launched the post-Renaissance idea that individual brain parts contributed to different functions.
The first truly accurate theory of cerebral localization appeared in the mid-1700s. Emanuel Swedenborg (1688–1772) postulated that different functions were represented in different anatomical loci within the cerebral cortex. He argued that the variation in clinical signs observed from individuals with brain trauma could only be explained by anatomical separation of function. He identified distinct cerebral regions separated by fissures and gyri. He placed the motor cortex in the anterior portion of the brain and further identified a somatotopic representation by which the muscles of the extremities were controlled by upper convolutions, the trunk by the middle convolutions, and the neck and head by the lower convolutions. Unfortunately for him, his work was not widely distributed until after his death. While Swedenborg was developing his ideas of cerebral localization, he “began to experience mystical visions” (Finger 1994, p. 30), which led him to abandon his work in the neurological sciences in favor of a religious following.
The modern era of cerebral localization and functional specificity began in the 1800s with the writings of Bell (1774–1842), Bouillaud (1796–1881), Andral (1797–1846), and Broca (1824–1880). Much of the early work was in reaction against phrenology, the concept promoted by Franz Joseph Gall and his devoted follower Johann Spurzheim (1776–1832). Phrenology is the pseudoscience of attributing brain function with structure primarily by examining the surface of the cranium. Franz Joseph Gall collected over 300 skulls of individuals from the extremes of society (scholars, statesmen, criminals, lunatics) and attempted to correlate mental characteristics with cranial surface maps. Phrenology was adopted by physicians of the time for diagnosing neurological disease, by criminologists for attributing criminal behavior to a physically defective brain, and by scholars for selecting individuals with particular intellectual or artistic talents.
Phrenology remained popular throughout Europe and the Unites States from the 1780s until Gall died in 1828. Debate dominated the scientific community throughout the 1800s with conservatives following the lead of Marie-Jean Flourens (1774–1867), who advocated greater emphasis on laboratory and animal study, while liberal followers of Aubertin (1825–1893) and others advocated localization based on clinical evidence. Conservatives cautioned against the direct structure–function theory on the basis of inconsistent laboratory studies. Localizationalists, on the other hand, advanced series after series of clinical cases supporting specific functions (e.g., speech or memory) to autopsy confirmed cortical regions. The two camps merged with experimental confirmation of Broca’s clinical report of functional localization of motor behavior.
In 1869, Eduard Hitzig (1838–1907) and Gustav Fritsch (1838–1927) conducted an experiment (on a dog) proving that cortical localization need not be limited to a single function. Hitzig and Fritsch’s experiment confirmed that applying electrical current to the frontal cortex in close proximity to the Broca’s motor speech area impaired motor function. Hitzig and Fritsch replicated their finding in other animals and found distinctive cortical sites that elicited motor responses throughout the extremities, neck, and head on the opposite side of the stimulation. Further mapping led to unequivocal support for the existence of the motor cortex. Sir David Ferrier (1843–1928) replicated Hitzig and Fritsch’s work and extended it to the monkey brain. Using more precise electrical stimulation and careful mapping, Ferrier was able to map a region of the motor cortex that corresponded to movement of a single finger. Ferrier’s work was summarized in an 1876 publication entitled The Functions of the Brain that led neurosurgeons at the time to rely on functional maps for guidance during surgery.
Perhaps the most successful example of cortical mapping from the 1800s was the numbering system published by Korbinian Brodmann in 1909. His map of 52 discrete cortical regions was based on differences in structural and cellular composition. The map clearly distinguished motor from sensory areas. The map accurately delineated regions with fine granularity despite variation in experimental methods and across species. The histological delineation of cortical areas inevitably corresponded to functional specificity. Today, modern neurosurgery relies on many of the same principles of cortical mapping pioneered by the localizationalists throughout the 1800s.
In the late 1800s, Sigmund Exner (1846–1926) published a book entitled Untersuchungen über die Lokalisation der Functionen in der Grosshirnrinde des Menschen (Studies on the Localization of Functions in the Cerebral Cortex of Humans). In his book, Exner (1881) described a specific area of the cerebral cortex (the posterior part of the left middle frontal gyrus), which he attributed to handwriting. This area rapidly became known as Exner’s “writing center.” Based on just a few cases, Exner claimed that the posterior portion left middle frontal gyrus was the writing equivalent of Broca’s motor speech area.
Despite having sparse data to back up this claim, the notion of a specific writing center ignited passionate debate throughout the scientific community (Roux et al. 2010). In his book, Exner described only four cases with agraphia that he associated with lesions to this area. Unfortunately, closer inspection revealed that in only one of these cases was the agraphia not accompanied by either hemiparesis or aphasia, which would lead to writing difficulties for reasons other than execution of the handwriting motor program (e.g., muscle weakness or paralysis or expressive language impairment).
As we will see later in this chapter, modern science has failed to support the notion of a single localized writing center. Rather, this “writing center” is likely to involve a network of cortical areas. Nonetheless, modern writers continue to refer to Exner’s area as a writing center (e.g., Seitz et al. 1997; Sugihara, Kaminaga, and Sugishita 2006). However, because of its role in language processing, the posterior portion of the left middle frontal gyrus is not likely to be a member of this putative network.
Neuroanatomical Bases of Hand Motor Control
Functional Organization
Prior to undertaking a discussion of the anatomical regions, pathways, and circuits underlying hand motor control, it is important to understand the general organizational structure of the human nervous system. By understanding the fundamental organization of the brain, we can formulate hypotheses or predictions about what to expect in the form of altered handwriting following localized injury to the brain. In this section, we will introduce several approaches to understanding how the brain is organized for hand movement. Most of the organizational schemes hold that motor functions can be either spatially or topographically mapped onto a given brain region or network. Other organizational schemes rely on a hierarchical approach whereby certain brain areas (e.g., cortex) exhibit high-level, integrative or executive functions while other areas (e.g., basal ganglia) function in fine-tuning.
From the perspective of gross neuroanatomy, the nervous system can be divided into the central nervous system (CNS), consisting of the brain and spinal cord, and the peripheral nervous system (PNS), consisting of the nerves running to and from the spinal cord and periphery (i.e., muscle). Within the CNS, brain functions may be further organized into anterior–posterior or left–right dimensions. At this level, motor functions are typically attributed to anterior regions, while sensory processes are attributed to posterior regions. Cortical representation of the musculoskeletal system is bilateral. That is, sensory-motor functions of the left side of the body are regulated, at least at the cortical level, by the contralateral or right cerebral cortex, whereas sensory-motor functions of the right side of the body are regulated by the left cerebral cortex. This lateralization is well preserved throughout the cortex and spinal cord. Figure 1.2 shows the four main lobes of the left cerebral cortex including the frontal, parietal, temporal, and occipital cortices. Demarcation boundaries are based on Brodmann’s cytoarchitectonic maps (see Figure 1.4 later in the chapter)
Another approach to understanding the functional organization of the CNS is based on the principle that the representation throughout the CNS is topographically organized. That is, the brain’s functional organization for a given body area (e.g., the hand) is represented by a spatial map that is preserved throughout the brain’s vertical hierarchy from the cortex through the basal ganglia and the brain stem and into the spinal column. This topographic representation generally holds that representation of lower extremities is topographically represented toward midline regions of the brain and, as we move from midline to lateral regions of the brain, representation follows from upper extremities to head and face, respectively. This scheme is commonly portrayed as a homunculus, or a representation of an anatomically deformed human being mapped onto the brain surface. Such a homunculus is depicted in Figure 1.3.
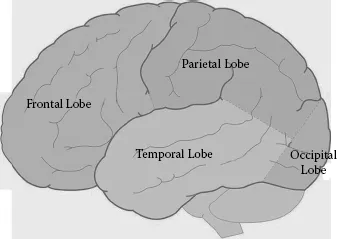
Figure 1.2 Demarcation boundaries are based on Brodmann’s cytoarchitectonic maps.
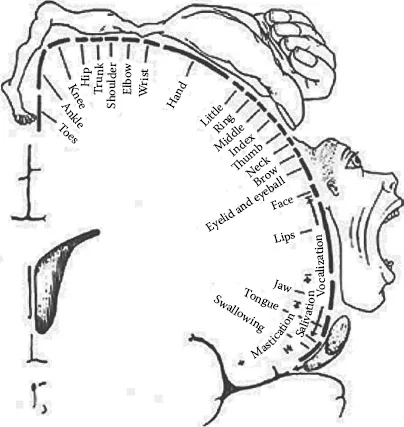
Figure 1.3 Drawing of the motor homunculus. The homunculus depicts a model of the human brain to illustrate anatomical representation of body movement. (From Penfield, W., and Rasmussen, T. 1950. The Cerebral Cortex of Man: A Clinical Study of Localization of Function. New York: MacMillan. Copyright Gale, a part of Cengage Learning, Inc. Reproduced by permission. www.cengage.com/permissions.)
The precise control of hand movement for handwriting involves multiple motor and sensory areas throughout the central and peripheral nervous systems. In the following sections, we review the available evidence for specific roles of the motor and association cortices, basal ganglia, cerebellum, brain stem, and spinal cord in the control of handwriting.
Motor and Association Cortices
Three cortical regions play key roles in hand motor control: the primary motor area (Brodmann area 4), the premotor area (Brodmann area 6), and the supplementary motor area (SMA; located midline to area 5). Figure 1.4 shows the demarcations of these areas in a diagram of the cortex. While the primary motor area and the premotor area extend from the medial to lateral portions of the cerebral hemisphere, the SMA extends into the mesial surface of the cerebral hemisphere within the longitudinal fissure that separates the two hemispheres (not shown in the diagram).
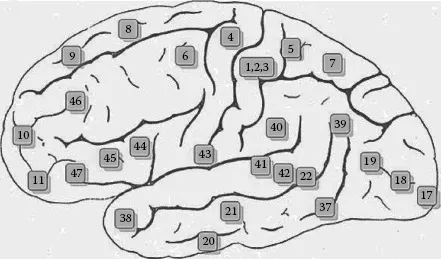
Figure 1.4 Broadm...
Table of contents
- Cover
- Title Page
- Copyright Page
- Contents
- Series Preface—International Forensic Science Series
- Foreword
- Preface
- Acknowledgments
- The Authors
- Section I: FUNDAMENTAL ASPECTS OF MOTOR CONTROL AND HANDWRITING
- Section II: KINEMATICS OF SIGNATURE AUTHENTICATION
- Section III: NEUROLOGIC DISEASE, DRUGS, AND THE EFFECTS OF AGING
- Bibliography
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Neuroscience of Handwriting by Michael P. Caligiuri,Linton A. Mohammed in PDF and/or ePUB format, as well as other popular books in Law & Forensic Science. We have over 1.5 million books available in our catalogue for you to explore.