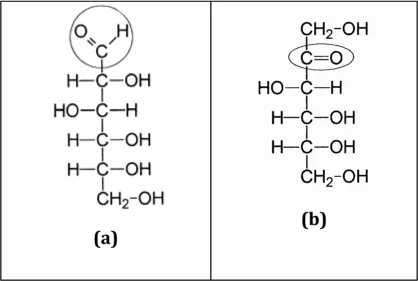
Basically, two different systematic approaches can be used for a classification of carbohydrates: as carbohydrates either contain hydroxy and aldehyde functions (see Fig. 1.1a, glucose) or, alternatively, hydroxy and ketone functions (Fig. 1.1b, fructose), this discrimination may form a basis for a classification. Another approach that is often being used in basic monographs is based on the systematic discrimination between monosaccharides, disaccharides, oligosaccharides and polysaccharides. In the present chapter, both approaches will be combined by including the functional aspects in the sub-chapter mono-saccharides. As the terms suggest, mono- and di-saccharides consist of one and two sugar units, respectively, while oligosaccharides are assumed to contain 3 to 10 subunits linked together (according to the Greek word oligos, for “a few”). Polysaccharides contain more than 10 subunits.
1.2.1 Monosaccharides
The early constitutional as well as stereochemical investigations are closely related to the names Emil Fischer, Kiliani, and Tollens (Lichtenthaler, 1992). It was inferred from these basic results that monosaccharides can be considered to be oxidation products of alcohols that contain several hydroxy groups, resulting in polyhydroxy aldehydes or polyhydroxy ketones. The former group of oxidation products is nominated aldoses, while the latter group is called ketoses.
Figure 1.1 (a) Glucose and (b) fructose.
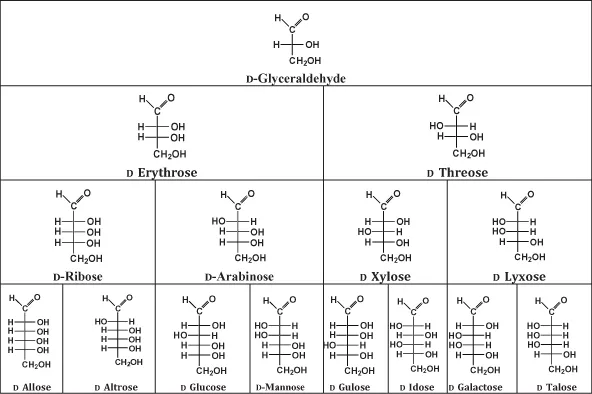
On the basis of the above definitions the so-called sugar tree can be derived, starting with the most simple compound that fulfils these characteristics, the aldotriose 2,3-dihydroxy propanal (“glyceraldehyde”). Insertion of an additional H–C–OH group leads to aldotetroses, further insertion of another H–C–OH group results in aldopentoses, and finally, addition of another H–C–OH group supplies the aldohexoses. This procedure is depicted in Fig. 1.2.
Figure 1.2 “Sugar tree” showing the stereoisomers of aldotetroses, aldopentoses, and aldohexoses as systematically derived from D-glyceraldehyde.
1.2.1.1 Configuration and nomenclature
A full understanding of the sugar tree requires information on the absolute structures of these monosaccharides. The basic compound glyceraldehyde contains one stereogenic centre at the C2 atom. Accordingly two enantiomers can be defined. In order to discriminate between these enantiomers, different notations can be used. In Fig. 1.3 the so-called Fischer projection is displayed that follows very strict drawing rules: the stereogenic centre is placed in the centre of two crossing lines. Please note that the atom in this centre must not appear, i.e., in the case of the chiral glyceraldehyde the C2 must not be drawn. Otherwise a constitutional formula without any stereochemical information would be obtained. Furthermore, the carbon with the highest oxidation number, in the case of the monosaccharides an aldehyde or keto function, appears at the top or as close to the top as possible. The carbon chain backbone is being written in the vertical line. If the hydroxy group at the stereogenic centre appears at the right hand side, the D-enantiomer is obtained, and if the hydroxy group is placed at the left hand side, the L-enantiomer is assigned. This means that in the sugar tree shown in Fig. 1.2 exclusively the D-glyceraldehyde is given. Furthermore, it should be noted that IUPAC rules require the D and L prefixes to be written in small caps.
Figure 1.3 Fischer projection of the chiral glyceraldehyde.
The unequivocal advantage of the Fischer projection is that the absolute structure can be inferred, according to the following definition: the horizontal lines of the central cross lines are pointing towards the observer, while the two vertical lines are directing into the plane. Addition of an additional H–C–OH group (aldotetroses) gives rise to a second stereogenic centre. As a consequence, 2n = 4 stereoisomers can be expected, i.e., two pairs of enantiomers, where the stereochemical relation between these two pairs is denominated with the term diastereomer. It is worth noting that enantiomers basically exhibit the same physical parameters, while diastereomers may show different ones, provided that no enantioselective interactions with other chiral molecules or polarized light is encountered. In the case of aldopentoses, three stereogenic centres are present, and for aldohexoses four stereogenic centres are found, resulting in 23 = 8 and 24 = 16 stereoisomers, respectively. For example, in the case of the aldohexoses the eight stereoisomers shown in Fig. 1.2 plus the respective mirror images (enantiomers) exist. Basically, at each stereogenic centre a D- or L- notation is conceivable. However, in order to facilitate the nomenclature, a clear definition allows an assignment to the D- and L- notation: the stereogenic centre that is most remote of the carbonyl function (carbon exhibiting the highest oxidation number) determines the assignment to the group of D- and L-stereoisomers. However, it has to be noted that the absolute structure of the molecule cannot be directly inferred from the name; it can only be indirectly obtained on the background of additional knowledge of the glucose structure. Diastereoisomers that exhibit the opposite configuration at only one of two or more tetrahedral stereogenic centres present in the respective molecular entities are called epimers. This holds, for example, for the diastereomeric pair D-glucose and D-mannose (see Fig. 1.2).
Alternatively, the R-/S- notation that is based on the Cahn-Ingold-Prelog (CIP) rules can be applied to carbohydrates with several stereogenic centres. For example, strict assignment of each stereogenic centre in D-glucose to the R-/S- notation results in the exact IUPAC name (2R,3S,4R,5R,6)-pentahydroxyhexanal. In contrast to the D/L notation, this name allows an exact assignment of each stereogenic centre to its absolute structure. However, as this name is very inconvenient in publications, the name D-glucose as well as the respective names of other sugars according to the D/L notation are still dominating.
1.2.1.2 Ring structures of carbohydrates
As early as 1883 Tollens had recognized that simple aldohexoses such as glucose do not show all characteristics of aldehydes. In particular, no addition of sodium hydrogensulfate or ammoniak were observed, and no reaction with Schiff's reagent takes place. Tollens inferred from these observations that simple sugars do not prefer the aldehyde or keto form arrangement. Instead, an intramolecular hemiacetal formation may lead to cyclic structures, as shown in Fig. 1.4. These Tollens formulae suggest that basically five- or six-membered rings can be formed by reaction of the carbonyl function with the hydroxy group at C4 or C5. Furthermore, it is evident that an additional stereogenic centre is formed at the C1 position. If the hydroxy group...