Plants depend on physiological mechanisms to combat adverse environmental conditions, such as pathogen attack, wounding, drought, cold, freezing, salt, UV, intense light, heavy metals and SO2. Many of these cause excess production of active oxygen species in plant cells. Plants have evolved complex defense systems against such oxidative stress. The
Atomic oxygen is the most abundant element in the earth’s crust; molecular oxygen in the atmosphere and water is required to support all forms of aerobic life. The present oxygen reservoir (37 Emol, 1 Emol=1018 moles) has built up as a result of photosynthesis, a process that liberates dioxygen from water. It is kept approximately constant by respiration, in which O2 is used as the ultimate electron acceptor. In addition, oxygen atoms are “fixed” into various organic molecules by a variety of enzymes (e.g. oxygenases) and non-enzymatic processes (Gilbert, 1981; Elstner, 1982, 1987). Aerobic organisms must, however, cope with the adverse effects of oxygen. At higher-thanatmospheric concentrations, dioxygen may inhibit or inactivate certain enzymes and it also competes with photosynthetic CO2 fixation by ribulose-1,5-bisphosphate carboxylase/ oxygenase, increasing the energetic cost of photosynthesis. Still, the toxic effect of oxygen is mainly exerted by its reactive derivatives, whereas ground-state dioxygen is rather unreactive and can peacefully co-exist with organic matter. This characteristic is explained by the parallel spins of two unpaired electrons of dioxygen, imposing an energetic barrier on its reaction with non-radical compounds (the “spin restriction”). In order to become chemically reactive, dioxygen must be physically or chemically activated (Table 1).
Physical activation occurs mainly by transfer of excitation energy from a photoactivated pigment such as an excited chlorophyll molecule to dioxygen. The latter absorbs sufficient energy and, as a result, the spin of one electron is inverted. The first singlet state of oxygen (designated 1O2 or 1 ΔgO2) is a prevalent reactive species. It is highly diffusible and capable of reacting with organic molecules (whose electrons are usually paired), and damaging photosynthetic membranes.
Chemical activation is the other mechanism to circumvent spin restriction. It occurs by univalent reduction of dioxygen, i.e. addition of electrons one by one. Four electrons (and four protons) are required for the full reduction of dioxygen to water; all three intermediates of univalent reduction, namely superoxide (
) hydrogen peroxide (H2O2), and the hydroxyl radical (OH•), are chemically reactive and biologically toxic (Elstner, 1987; Hamilton, 1991; McKersie and Leshem, 1994; Yu, 1994). This toxicity is reflected by their short half-lives before reacting with cellular components, as compared to that of dioxygen (>100 sec; Table 1). Reactive oxygen species colliding with an organic molecule may extract an electron from it, rendering it a radical capable of propagating a chain reaction, e.g. the peroxyl (ROO•) and alkoxyl (RO•) radicals.
Superoxide is the first reduction product of ground state-oxygen, capable of both oxidation and reduction. It may react to produce several other reactive species, and may undergo spontaneous or enzymatic dismutation to H2O2.
Hydrogen peroxide is not a free radical, but participates as oxidant or reductant in many cellular reactions. Unlike superoxide, H2O2 is highly diffusible through membranes and aqueous compartments and it may directly inactivate sensitive enzymes at a low
concentration. Much like superoxide, H2O2 is rather stable and therefore less toxic than other reactive oxygen species; the main threat imposed by both superoxide and H2O2 lies in their ability to generate highly reactive hydroxyl radicals.
Table 1. Formation and characteristics (compared to molecular oxygen) of major reactive oxygen species
The hydroxyl radical is the most powerful oxidizing species in biological systems. It will react non-specifically with any biological molecule, and this will limit its diffusion within the cell to a distance of two molecular diameters from its site of production. No specific scavengers of OH• are known, although several metabolites, such as urea or glucose, were proposed as hydroxyl scavengers in animal systems. Recently, a role for OH• in cell wall polysaccharide metabolism has been proposed (Fry, 1998).
The different reactive species described above will cause, to varying extents, (i) inhibition of sensitive enzymes (some specific examples are discussed below), (ii) chlorophyll degradation or “bleaching”, (iii) lipid peroxidation; free radicals, H2O2, and singlet oxygen readily attack unsaturated fatty acids, yielding lipid hydroperoxides, and, in the presence of metal catalysts, alkoxyl and peroxyl radicals that propagate chain reactions in the membranes, changing and disrupting lipid structure and membrane organization and integrity (Yu, 1994). In addition, some aldehydes and hydrocarbons produced by lipid peroxidation exert cytotoxic effects in animal systems (Esterbauer et al., 1990). (iv) Indiscriminate attack by hydroxyl radicals of organic molecules, including DNA. A variety of oxidatively altered DNA species can be identified following OH• attack, including base alterations and strand breaks that may be difficult to repair or tolerate (Kasai et al, 1986). Proteins exposed to OH• undergo typical modifications, including specific amino acid alterations, polypeptide fragmentation, aggregation, denaturation, and susceptibility to proteolysis (Wolff et al, 1986).
BIOLOGICAL SOURCES OF REACTIVE OXYGEN
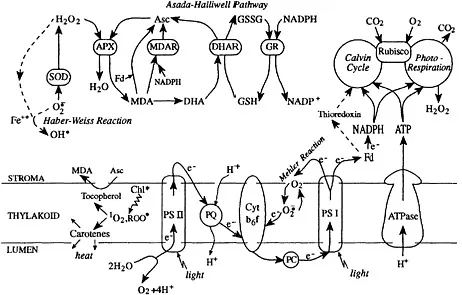
It is well-established that the formation of reactive oxygen species (ROS) accompanies normal metabolic processes in all aerobic organisms. We will describe in some detail the source of different reactive species in plant cells. Figure 1 illustrates many of the physiological pathways that are discussed along the chapter. However, many pioneer discoveries on oxygen radicals were made with a facultative aerobe, Escherichia coli (Fridovich, 1991). In aerobically grown E. coli,
is produced by reduction of dioxygen during membrane-associated electron transport. Only 0.04% of the electrons “leak”, but a mechanism to prevent superoxide accumulation is required. Superoxide levels were measured using
Figure 1. Biochemical chart of the main physiological processes involving reactive oxygen species in the chloroplast.
E. coli membrane preparations of mutant strains devoid of superoxide dismutase (sodA, sodB) (Carlioz and Touati, 1986; Imlay and Fridovich, 1991). Such mutants exhibit slow growth and several auxotrophies, demonstrating the vital role of SOD in decreasing steady state levels of
down to 2×10–10 M. It is interesting to note that one of the most superoxide-sensitive enzymes in this system is E, coli aconitase of the citric acid cycle (Gardner and Fridovich, 1991). The reversible oxidative inactivation of aconitase by
has been suggested to fulfill a defensive role of “circuit breaking” to cut off NADPH production, thus avoiding further build-up of superoxide by respiration. Examples of metabolic inactivation/deviation pathways used to avoid reducing conditions in the photosynthetic apparatus are discussed below.
Reactive Oxygen Formation in Plant Chloroplasts
Chloroplasts are the major source of reactive oxygen forms in plants: they harvest light energy at high efficiency, produce reducing equivalents, such as NADPH, and generate fluxes of dioxygen: indeed the most “radical-prone” conditions one could imagine. Several independent pathways, or sites of oxygen activation, have been described in chloroplasts, leading to the production of all of the above reactive species (Elstner, 1982; 1991; Asada, 1994; Foyer and Harbinson, 1994). The most important is the reducing side of photosystem I (PSI), where an electron may be passed from a membrane-bound carrier to O2 (the “Mehler reaction”), instead of flowing to downstream carriers that finally reduce NADP+ (Figure 1). Under conditions that limit the availability of electron acceptors from PSI, i.e., when the Calvin cycle does not consume NADPH rapidly enough, superoxide will form within the membrane. At low pH, superoxide may spontaneously dismutate to the more diffusible H2O2; otherwise it may interact with plastocyanine or cytochrome f and reduce them, resulting in a superoxide-mediated cyclic electron flow around PSI (Hormann et al, 1993). This mechanism actually suggests a regulatory role for superoxide production, namely to divert, or cycle, excessive flow of electrons and, at the same time, prevent the diffusion of radicals away from the membrane.
Hydrogen peroxide is mostly produced in chloroplasts by disproportionation of superoxide by SOD, which is much faster than spontaneous dismutation. Another source of H2O2 is photorespiration that is initiated by the oxygenase activity of ribulose-1,5- bisphosphate carboxylase in the chloroplast stroma, followed by the production of H2O2 in the peroxisomes (Figure 1). Photorespiration may be interpreted as a protective mechanism that recycles electron acceptors and allows photosynthetic electron flow to continue under conditions of low carbon fixation. Compared to electron-cycling around PSI (discussed above as a possible protective pathway), the photorespiratory cycle would dissipate both ATP and NADPH (Wu et al, 1991). An important mechanism of regulation that couples carbon fixation in the stroma to photosynthetic electron flow, is the reversible inactivation of Calvin cycle enzymes when electron carriers of the light reaction are oxidized, and their re-activation when the carriers are reduced. Such regulation is mediated by thioredoxin, stromal pH, and other factors (Figure 1; Foyer et al, 1992). Hydrogen peroxide will disrupt this delicate mechanism by oxidizing thiol groups and inactivating the Calvin cycle enzymes irreversibly; it must therefore be kept below micromolar concentrations in chloroplasts (Foyer and Harbinson, 1994). It will also inactivate copper/zinc (Cu/Zn) SOD.
Hydroxyl radicals may be formed in all living cells in a reaction catalyzed by the transition metal ions, iron and copper, when bot...
Table of contents
COVER PAGE
TITLE PAGE
COPYRIGHT PAGE
PREFACE
CONTRIBUTORS
ABBREVIATIONS
1. OXIDATIVE STRESS: AN INTRODUCTION
2. THE CONTRIBUTION OF PHOTOSYNTHETIC OXYGEN METABOLISM TO OXIDATIVE STRESS IN PLANTS
3. LOW-TEMPERATURE STRESS AND ANTIOXIDANT DEFENSE MECHANISMS IN HIGHER PLANTS
4. OXIDATIVE STRESS AND DEFENSE REACTIONS IN PLANTS EXPOSED TO AIR POLLUTANTS AND UV-B RADIATION
5. OXIDATIVE BURST AND THE ROLE OF REACTIVE OXYGEN SPECIES IN PLANT-PATHOGEN INTERACTIONS
6. PHOTOSENSITIZING TETRAPYRROLES INDUCE ANTIOXIDATIVE AND PATHOGEN DEFENSE RESPONSES IN PLANTS
7. METAL ION-ACTIVATED OXIDATIVE STRESS AND ITS CONTROL
8. ENGINEERING STRESS TOLERANCE IN MAIZE
9. EARLY EVENTS IN ENVIRONMENTAL STRESSES IN PLANTS—INDUCTION MECHANISMS OF OXIDATIVE STRESS
10. THE MOLECULAR BIOLOGY OF THE ASCORBATE- GLUTATHIONE CYCLE IN HIGHER PLANTS
11. ASCORBATE METABOLISM AND STRESS
12. GLUTATHIONE BIOSYNTHESIS IN PLANTS
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Oxidative Stress in Plants by Dirk Inze,Marc Van Montagu in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biochemistry. We have over 1.5 million books available in our catalogue for you to explore.