
- 214 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Introduction to Clinical Reproductive Endocrinology
About this book
Introduction to Clinical Reproductive Endocrinology covers the pathophysiological basis and management of clinical conditions related to reproductive endocrinology. The book describes the normal development and physiology of the hypothalamic-pituitary-gonadal axes and the structure and function of the hypothalamic, pituitary and gonadal hormones. The text then discusses the events of ovulation and fertilization and the endometrial changes of the menstrual cycle. The etiology, diagnosis, and management of problems encountered in reproductive medicine, including those of delayed and precocious puberty, primary and secondary amenorrhoea, and oligomenorrhoea are also considered. The book further tackles the premenstrual syndrome; endometriosis; female and male subfertility; problems of early pregnancy; menorrhagia; and the menopause. The text concludes by looking into the indications for, and the side-effects of, drugs used in reproductive medicine. Reproductive endocrinologists, obstetricians, and gynecologists and medical students taking courses related to reproductive endocrinology will find the book useful.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Endocrinology & MetabolismChapter 1
The hypothalamus and the pituitary gland
Publisher Summary
This chapter discusses the hypothalamus and the pituitary gland vis-à-vis the reproductive function and gonadal development. It also discusses the hormones secreted by the hypothalamus and the pituitary gland. The hypothalamus and the pituitary gland play a vital role in the control of gonadal development and reproductive function. Peptide and glycoprotein trophic hormones—including hypothalamic releasing hormones and some of the anterior pituitary hormones—act by binding to specific receptors in the cell membranes on the surface of target cells. They do not enter the cells but stimulate intracellular activity by a variety of mechanisms. Several hypothalamic hormones are secreted directly from axon terminals in the median eminence into the highly innervated capillaries of the hypothalamus which drain into the portal vessels. The portal vessels supply the anterior lobe of the pituitary and thus deliver hypothalamic hormones to the anterior pituitary. It is possible that the blood also flows in the portal veins in a retrograde manner from the pituitary to the hypothalamus. The chapter presents the hypothalamic hormones one of which is the luteinizing hormone releasing hormone (LHRH). It is produced by hypothalamic neurones, particularly in the region of the arcuate nucleus, and secreted from axon terminals in the median eminence into the portal circulation in a pulsatile manner. TRH is a tripeptide which is produced in the hypothalamus, as well as in other parts of the brain, and is secreted into the portal vessels. It acts on cells of the anterior pituitary to stimulate the production of both thyroid stimulating hormone and prolactin.
The hypothalamus and the pituitary play a vital role in the control of gonadal development and reproductive function. Considerable advances have been made in the understanding of the physiology of the hypothalamic–pituitary axis but much remains to be elucidated. It is clear that there is a complex interplay involving neural and vascular connections between the hypothalamus and pituitary. Transection of these connections leads to cessation of secretion of most of the pituitary hormones. Thus pituitary function is in one sense largely under the control of the hypothalamus. However, the function of the hypothalamic–pituitary axis is in turn influenced by many other local and distant inputs, including both negative and positive feedback from target glands. The whole system is exceedingly complex and very finely balanced.
Hormones
Hormones were defined classically, by Starling, as chemical transmitters released from specialized cells into the bloodstream and carried thence to other responsive cells where they exert their effects.
It has been realized that the situation is much more complicated than this, that many cells (including those of unicellular organisms) can produce a variety of hormones, and that the formation of endocrine glands is a relatively late development in evolution. This explains why more and more hormones are being shown to be produced physiologically in unexpected sites and also pathologically by malignant cells.
Hormones provide a means of communication, not only at a distance but also locally between and within cells. The term paracrine effect indicates the response of a group of cells to a hormone which diffuses locally from its site of production in a neighbouring group of cells (Figure 1.1). An autocrine effect is the response of a cell to its own hormones.
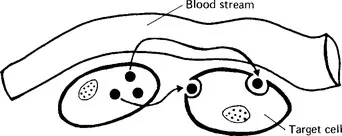
Figure 1.1 Paracrine (localized) hormone action
Mechanism of action of trophic hormones
Peptide and glycoprotein trophic hormones (including hypothalamic releasing hormones and some of the anterior pituitary hormones) act by binding to specific receptors in the cell membranes on the surface of target cells. They do not enter the cells but stimulate intracellular activity by a variety of mechanisms. Most of the polypeptide hormones stimulate the adenyl cyclase system causing the conversion of adenosine 5′-triphosphate to the second messenger cyclic adenosine 3′5′-monophosphate, which activates production of the relevant hormone. In the case of the hypothalamic releasing hormones other second messengers such as inositol 1,3,4-triphosphate and diacylglycerol are involved and there is an increase in intracellular calcium levels.
The hypothalamus
The structure of the hypothalamic–pituitary region is shown in diagrammatic form in Figure 1.2.
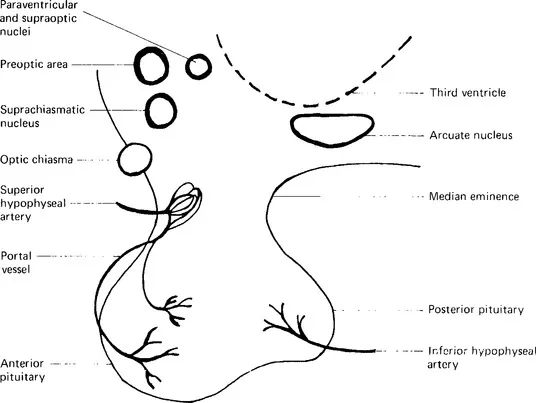
Figure 1.2 Diagrammatic representation of the hypothalamic-pituitary region
The hypothalamus is a small region of the brain, only weighing approximately 10 g. It lies at the base of the brain just above and posterior to the optic chiasma and is adjacent to the anterior part of the third ventricle. It consists of medial and lateral areas. The medial hypothalamic area is subdivided into a number of smaller regions, including the preoptic, paraventricular, supraoptic and anterior hypothalamic nuclei (in the anterior group), the ventromedial, dorsomedial, lateral tuberal and arcuate nuclei (in the tuberal group) and the posterior hypothalamic, mamillary, supramamillary and tuberomamillary nuclei (in the posterior group). Most of the neurones which produce hypothalamic hormones are thought to be in the tuberal area. The lateral hypothalamic area lies lateral to the medial hypothalamic area; it contains neurones which connect the rest of the brain with the medial hypothalamic nuclei.
Neurones are highly differentiated cells. They consist of a cell body (containing the nucleus), an axon and a number of dendrites (Figure 1.3). Axons vary in length from being very short to up to a metre or more long. They end at synapses in a number of axon terminals. Communication at a synapse is achieved by release of chemical transmitters stored in vesicles in the presynaptic axon. Some neurotransmitters, such as the catecholamines, acetylcholine, serotonin and γ-aminobutyric acid (GABA), are produced in the nerve terminals, whereas others are produced in the cell body. Thus the cell bodies of neurones originating in the supraoptic and paraventricular nuclei produce peptides that are transported down their axons and released from the axon terminals; they also communicate with other neurones via conventional synapses.
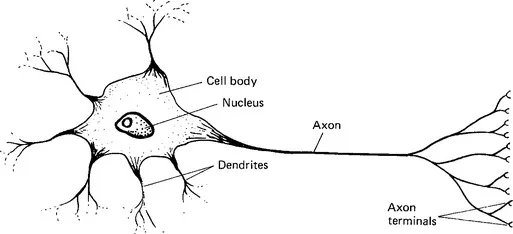
Figure 1.3 Neurone
Several hypothalamic hormones are secreted directly from axon terminals in the median eminence into the highly innervated capillaries of the hypothalamus which drain into the portal vessels. The portal vessels supply the anterior lobe of the pituitary and thus deliver hypothalamic hormones to the anterior pituitary.
It is possible that blood also flows in the portal veins in a retrograde manner from the pituitary to the hypothalamus (Bergland and Page, 1978). Substances can also pass from the cerebrospinal fluid to the portal plexus (and possibly in the opposite direction; Bergland and Page, 1978) via tanycytes, as well as being secreted directly into the cerebrospinal fluid by secretory neurones. Tanycytes are specialized ciliated ependymal cells whose cell bodies line the third ventricle; they extend down to the median eminence.
The term blood–brain barrier refers to the fact that most brain capillaries are less permeable than capillaries in the rest of the body, as they do not have fenestrations in their endothelial cells and because they are covered by astrocytic processes. These differences from other capillaries are not, however, found in the capillaries of the median eminence, which therefore lies outside the blood–brain barrier.
It has been shown that a large number of peptides are produced in the central nervous system (CNS). Their production may be widespread, as in the case of thyrotropin releasing hormone (TRH) and the opioid peptides, or localized. Their presence in particular sites can be mapped by immunocytochemical techniques using specific antibodies. Several peptide hormones are initially produced in the form of a prohormone that is then broken down into smaller units, as in the case of adrenocorticotrophic hormone (ACTH), β-melanocyte stimulating hormone (β-MSH), β-endorphin and other peptides which are derived from proopiomelanocortin. Neuropeptides are produced not only in the CNS but also in many peripheral tissues, including the placenta.
Specific steroid receptors, which allow negative and positive feedback by steroid hormones, have been demonstrated in the pituitary and in several areas of the hypothalamus, as well as in other parts of the brain, using labelled steroids.
Hypothalamic hormones
Luteinizing hormone releasing hormone (LHRH)
LHRH is a decapeptide (Figure 1.4). It was isolated and synthesized in the United States in 1971. It is produced by hypothalamic neurones, particularly in the region of the arcuate nucleus, and secreted from axon terminals in the median eminence into the portal circulati...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Foreword
- Preface
- Chapter 1: The hypothalamus and the pituitary gland
- Chapter 2: Steroid hormones
- Chapter 3: The ovaries and the menstrual cycle; hormonal changes in pregnancy
- Chapter 4: Sexual differentiation
- Chapter 5: Puberty
- Chapter 6: Amenorrhoea
- Chapter 7: Oligomenorrhoea and hirsutism; the polycystic ovary syndrome
- Chapter 8: Premenstrual (postovulatory) syndrome
- Chapter 9: Endometriosis and adenomyosis
- Chapter 10: Female subfertility
- Chapter 11: Male subfertility
- Chapter 12: Miscarriage, ectopic pregnancy and hydatidiform mole
- Chapter 13: Menorrhagia and dysmenorrhoea
- Chapter 14: The menopause
- Chapter 15: Drug treatment
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Introduction to Clinical Reproductive Endocrinology by Gillian C. L. Lachelin in PDF and/or ePUB format, as well as other popular books in Medicine & Endocrinology & Metabolism. We have over 1.5 million books available in our catalogue for you to explore.