
eBook - ePub
The Physiological Basis of Rehabilitation Medicine
- 782 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
The Physiological Basis of Rehabilitation Medicine
About this book
The Physiological Basis of Rehabilitation Medicine: Second Edition presents a comprehensive examination of the management of patients with functional impairments due to disease or trauma. It discusses the distinction between disabilities and impairments per se. It addresses the method in which the human body adapts and compensates for the stress produced by physical injuries. Some of the topics covered in the book are the physiology of cerebellum and basal ganglia; description of upper and lower motor neurons; anatomy of the vascular supply to the brain; characteristics of the autonomic nervous system; structure, chemistry, and function of skeletal muscle; the receptors in muscle; and cardiopulmonary physiology. The role of muscle spindles in perception of limb position and movement is fully covered. An in-depth account of the physiology of synovial joints and articular cartilage are provided. The cellular and glandular components of the skin are completely presented. A chapter is devoted to the factors involve in wound healing. Another section focuses on the nerve conduction and neuromuscular transmission. The book can provide useful information to doctors, dermatologists, students, and researchers.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Upper and Lower Motor Neurons
MALCOLM B. CARPENTER
This work was supported by research grants CO7005 from the Department of Defense, Uniformed Services University of the Health Sciences, and NS-26658 from the National Institutes of Health, Bethesda, Maryland. The opinions and assertions contained herein are the private ones of the author and are not to be construed as official or reflecting the views of the Department of Defense or the Uniformed Services University of the Health Sciences. Experiments reported herein were conducted according to the principles set forth in the “Guide for the Care and Use of Laboratory Animals,” Institute of Laboratory Animal Resources, National Research Council, NIH Pub. No. 80-23.
Publisher Summary
This chapter presents an overview of the upper and lower motor neurons. Anterior horn cells and their peripheral processes (axons), which innervate striated muscle, constitute anatomic and physiologic units referred to as the final common motor pathway or the lower motor neuron. The concept of the lower motor neuron is not limited to spinal cord. Motor cranial nerve nuclei, which innervate muscles in the head and neck, also are classified as lower motor neurons. Anterior horn cells, which are the prototype for all motor neurons, lie in cell columns in the anterior gray horn of the spinal cord. Several distinct cell columns are evident in the anterior horn. A medial cell column extending throughout the length of the spinal cord, which is divisible into cell groups, innervates the long and short axial muscles. The lateral cell column innervates the remaining body musculature. In the thoracic region, the lateral cell column is small and innervates the intercostal and anterolateral trunk musculature. All the descending fibers systems that can influence or modify activities of the lower motor neuron constitute the upper motor neuron.
Loss of motor function in parts of the body owing to a neural lesion is a distressing and fearful event for anyone. A lesion involving the motor systems is evidenced by loss of voluntary movement, muscle weakness, loss of muscle tone, loss or alteration of reflex activity, abnormal postures, and ultimately substitution of inferior and awkward motor activity. Evaluation of loss or disturbances of motor function should begin by determining the site of the lesion. The location of the lesion frequently provides clues to its nature and insights into the specific pathology. Because disturbances of voluntary motor function may involve either the upper or lower motor neuron, the first step is to distinguish which is involved. This relatively simple, yet frequently puzzling, distinction is one of the cornerstones of clinical neurology.
The Lower Motor Neuron
Anterior horn cells and their peripheral processes (axons), which innervate striated muscle, constitute anatomic and physiologic units referred to as the final common motor pathway, or the lower motor neuron. The concept of the lower motor neuron is not limited to spinal cord. Motor cranial nerve nuclei, which innervate muscles in the head and neck, also are classified as lower motor neurons.
Anterior horn cells, the prototype for all motor neurons, lie in cell columns in the anterior gray horn of the spinal cord. Several distinct cell columns are evident in the anterior horn. A medial cell column extending throughout the length of the spinal cord, divisible into cell groups, innervates the long and short axial muscles. The lateral cell column innervates the remaining body musculature. In the thoracic region the lateral cell column is small and innervates the intercostal and anterolateral trunk musculature. In the cervical and lumbosacral enlargements the lateral cell column enlarges and consists of several large subgroups. Cell groups of the lateral cell column in the cord enlargements innervate the muscles of the extremities. Cells of the lateral column, located anteriorly and peripherally, innervate extensor and abductor muscle groups; cells located dorsal and central to these innervate flexor and adductor muscle groups (Figure 1-1). The spinal gray matter has cytoarchitectural lamination that divides it into separate zones.1,2 Anterior horn cells lie with Rexed’s lamina IX, characterized by large motor neurons, 30 to 100 μ in diameter (Figure 1-2). These large multipolar neurons have coarse Nissl granules, large central nuclei, and multiple dendrites that extend beyond the limits of lamina IX. Axons of these cells emerge via the ventral root and become mixed with dorsal root fibers distal to the dorsal root ganglion. Spinal nerves containing both motor and sensory fibers are referred to as mixed spinal nerves. Fibers of the mixed spinal nerve divide into dorsal and ventral primary rami (Figure 1-3). In the spinal enlargements the primary rami participate in plexus formation, resulting in the formation of the brachial and lumbosacral plexuses. Nerves given off from these plexuses provide innervation for the muscles of the upper and lower extremities.
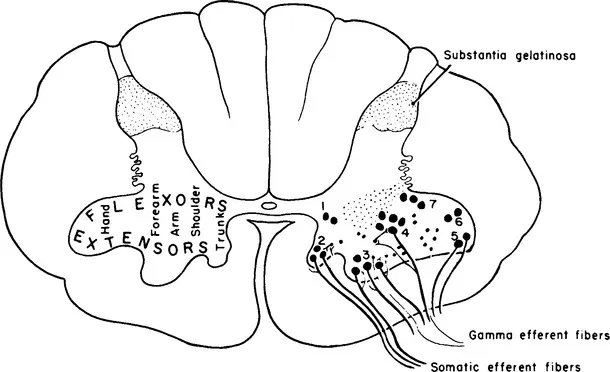
Figure 1-1 Diagram of the motor nuclei of the anterior gray horn in a lower cervical spinal segment. On the left, the approximate locations of neurons innervating different muscle groups are shown. On the right, groups of motor neurons are indicated by numbers. Both alpha and gamma fibers are shown emerging from the anterior horn. (From Carpenter MB. Core Text of Neuroanatomy, ed 3. Baltimore: Williams & Wilkins, 1985.)
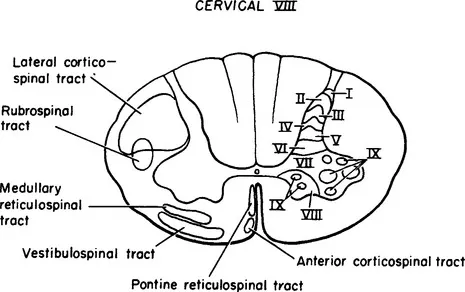
Figure 1-2 Drawing of a transverse section of the spinal cord at C-8 with the laminae of Rexed on the right and the position of the principal descending tracts indicated on the left.
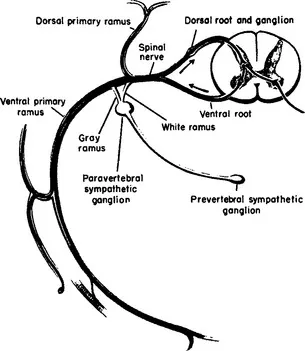
Figure 1-3 Schematic diagram of a thoracic spinal nerve showing peripheral branches and central connections. (From Noback CR, Demarest RJ. The Human Nervous System, ed 3. New York: McGraw-Hill, 1981.)
Not all cells in the anterior gray horn innervate striated muscle. Some, usually smaller than motor neurons, have processes confined to the spinal cord. These cells, referred to as internuncial neurons, have axons that project to other segments, to the opposite side of the spinal cord, or to motor neurons. Cells whose axons emerge from the spinal cord are known as root cells; these cells subserve effector functions. Root cells of the anterior horn are of two types: (1) alpha (α) motor neurons give rise to large fibers that innervate striatal (extrafusal) muscle and (2) gamma (γ) motor neurons innervate muscle spindles (intrafusal; Figures 1-2 and 1-4). The spectrum of myelinated fibers in the ventral roots indicates two groups of fibers. Approximately 70% of the fibers are between 8 and 13 μm in diameter and are classified as alpha fibers; the remaining 30% of the fibers, 3 to 8 μm in diameter, are designated gamma fibers. In addition, the ventral roots in thoracic and upper lumbar spinal segments contain unmyelinated or poorly myelinated preganglionic sympathetic fibers. Sacral ventral roots (S-2, S-3, and S-4) contain similar poorly myelinated preganglionic parasympathetic fibers. These preganglionic fibers project to various autonomic ganglia.
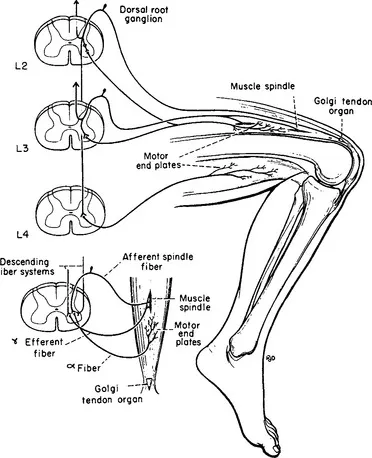
Figure 1-4 Schematic diagram of the sensory and motor elements involved in the patellar tendon reflex. Muscle spindle afferents are shown entering only the L-3 spinal segment; Golgi tendon organ afferents are shown entering only the L-2 segment. In this monosynaptic reflex afferent fibers enter L-2, L-3, and L-4 spinal segments and efferent fibers from the anterior horn cells at these same levels project to the extrafusal muscle fibers of the quadriceps femoris. Efferent fibers from L-4 projecting to the hamstrings represent part of the pathway involved in reciprocal inhibition. The small diagram on the left illustrates the gamma loop. Contractions of the polar parts of the muscle spindle initiate an afferent volley conducted centrally to alpha motor neurons. Discharge of the alpha motor neuron is conveyed to the motor end-plate of the same muscle. The gamma efferent fiber controls the sensitivity of the muscle spindle. (From Carpenter MB. Core Text of Neuroanatomy, ed 3. Baltimore: Williams & Wilkins, 1985.)
Alpha motor neurons are cholinergic and terminate upon skeletal muscle fibers in small, flattened expansions known as motor end-plates, which constitute the so-called myoneural junction. Electrical stimulation of a motor nerve causes quanta of acetylcholine (ACh) to be liberated at the myoneural junction, which produces contractions of muscle fibers. Following the contractions, acetylcholinesterase (AChE) hydrolyzes the ACh. Stimulation of a muscle nerve with graded shocks results in twitches of the muscle related directly to the strength of the stimulus, until the alpha spike reaches its full potential. Increasing the size of the stimulus does not produce a stronger contraction, even though gamma fibers may be discharged. Thus, impulses conducted by alpha motor fibers are related to the contractile elements of striated muscle and gamma motor neurons do not contribute directly to muscle contraction. Gamma fibers are distributed to the polar (contractile) portions of the muscle spindles. Contraction of the polar portions of the muscle spindle may be sufficient to cause the discharge of muscle spindle afferent fibers (group IA), but these contractions do not directly alter muscle tension or length. Impulses conducted by group IA enter the spinal cord via the dorsal root and distribute collaterals directly on alpha motor neurons. This two neuron linkage (one sensory neuron in the dorsal root ganglion and one motor neuron in the ventral horn) establishes part of the so-called gamma loop. The loop is closed by gamma efferent fibers, which arise from cells in the anterior horn and pass directly to polar parts of the muscle spindle (see Figure 1-4). Thus, impulses conveyed by gamma motor neurons can indirectly excite alpha motor neurons by causing the muscle spindle to fire. Part of this mechanism forms the basis for the myotatic (or stretch) reflex. Only part of the afferents from the muscle spindle pass to the anterior horn; a large part of the afferent volley ascends via relays in the spinal cord to the cerebellum.
The myotatic or deep tendon reflex is a monosynaptic reflex dependent on two neurons: one neuron in the dorsal root ganglion that receives afferent impulses from the muscle spindle and the alpha motor neuron that innervates the striated muscle containing the mus...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Dedication
- Contributing Authors
- Preface
- Acknowledgments
- Chapter 1: Upper and Lower Motor Neurons
- Chapter 2: Anatomy and Physiology of the Vascular Supply to the Brain
- Chapter 3: Cerebellum and Basal Ganglia
- Chapter 4: The Autonomic Nervous System
- Chapter 5: Skeletal Muscle: Structure, Chemistry, and Function
- Chapter 6: Receptors in Muscle and Their Role in Motor Control
- Chapter 7: Cardiopulmonary Physiology
- Chapter 8: Physiology of Synovial Joints and Articular Cartilage
- Chapter 9: Physiology of the Skin
- Chapter 10: Nerve Conduction and Neuromuscular Transmission
- Chapter 11: The Motor Unit and Muscle Action Potentials
- Chapter 12: Evoked Potentials
- Visual Evoked Potentials
- Auditory Evoked Potentials
- Somatosensory and Motor Evoked Potentials
- Chapter 13: Human Thermoregulation
- Chapter 14: Control of the Circulation in the Limbs
- Chapter 15: Exercise and Fatigue
- Chapter 16: Energy Expenditure During Ambulation
- Chapter 17: Physiologic Changes Associated with Bed Rest and Major Body Injury
- Chapter 18: Obesity and Weight Control
- Chapter 19: Urogenital Physiology
- Chapter 20: Autonomic Function in the Isolated Spinal Cord
- Chapter 21: Peripheral Nerve Regeneration
- Chapter 22: Biofeedback
- Chapter 23: The Physiologic Aspects and Clinical Application of Functional Electrical Stimulation in Rehabilitation
- Chapter 24: Central Nervous System Plasticity and Cognitive Remediation
- Chapter 25: Aging of the Reproductive System in Women: Menopause
- Chapter 26: Aphasia, Apraxia, and Agnosia
- Chapter 27: Skeletal Physiology and Osteoporosis
- Chapter 28: Biology of Aging in Humans
- Chapter 29: Pain and Suffering
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Physiological Basis of Rehabilitation Medicine by John A. Downey,Stanley J. Myers,Erwin G. Gonzalez in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Physiology. We have over 1.5 million books available in our catalogue for you to explore.