No matter what we do, however kind or generous our deeds may seem, a hidden motive of selfishness lurks--or so science has claimed for years. This book, whose publication promises to be a major scientific event, tells us differently. In Unto Others philosopher Elliott Sober and biologist David Sloan Wilson demonstrate once and for all that unselfish behavior is in fact an important feature of both biological and human nature. Their book provides a panoramic view of altruism throughout the animal kingdom--from self-sacrificing parasites to insects that subsume themselves in the superorganism of a colony to the human capacity for selflessness--even as it explains the evolutionary sense of such behavior.
Explaining how altruistic behavior can evolve by natural selection, this book finally gives credence to the idea of group selection that was originally proposed by Darwin but denounced as heretical in the 1960s. With their account of this controversy, Sober and Wilson offer a detailed case study of scientific change as well as an indisputable argument for group selection as a legitimate theory in evolutionary biology.
Unto Others also takes a novel evolutionary approach in explaining the ultimate psychological motives behind unselfish human behavior. Developing a theory of the proximate mechanisms that most likely evolved to motivate adaptive helping behavior, Sober and Wilson show how people and perhaps other species evolved the capacity to care for others as a goal in itself.
A truly interdisciplinary work that blends biology, philosophy, psychology, and anthropology, this book will permanently change not just our view of selfless behavior but also our understanding of many issues in evolutionary biology and the social sciences.

eBook - ePub
Unto Others
The Evolution and Psychology of Unselfish Behavior
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Unto Others
The Evolution and Psychology of Unselfish Behavior
About this book
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Publisher
Harvard University PressYear
1999Print ISBN
9780674930476
9780674930469
eBook ISBN
9780674256804
Topic
Scienze biologicheSubtopic
Psicologia socialeI
EVOLUTIONARY ALTRUISM
1
Altruism as a Biological Concept
The concept of altruism in everyday language seems to require an element of both action and motive. People who never help others are seldom considered altruists. On the other hand, when people do help others, we want to know why before we call them altruistic. Even saints could be regarded as selfish if they perceived their lives of sacrifice as tickets to heaven.
Despite the importance of motives in conventional definitions, evolutionary biologists define altruism entirely in terms of survival and reproduction. A behavior is altruistic when it increases the fitness of others and decreases the fitness of the actor. The challenge for the evolutionary biologist is to show how such self-sacrificial behaviors can evolve, regardless of how or even whether the individual thinks or feels as it performs the behavior.
It may seem strange that the biological concept of altruism can remain intuitive when the vital element of motives has been removed. Actually, the strangeness cuts both ways. Philosophical and psychological discussions of altruism often concentrate so heavily on motives that the actual act of helping is ignored. We intend to pay equal attention to actions and motives in this book, but we must begin by separating them. Our first task is to show how behaviors that benefit others at the expense of self can evolve. Our second task is to understand the psychological mechanisms that evolved to motivate these adaptive behaviors.
To see why evolutionists cannot resist talking about altruism, consider the trematode parasite Dicrocoelium dendriticum, which spends the adult stage of its life cycle in the liver of cows and sheep (Wickler 1976). The eggs exit with the feces of the mammalian host and are eaten by land snails, which serve as hosts for an asexual stage of the parasite life cycle. Two generations are spent within the snail before the parasite forms yet another stage, the cercaria, which exits the snail enveloped in a mucus mass that is ingested by ants. About fifty cercariae enter the ant along with its meal. Once inside, the parasites bore through the stomach wall and one of them migrates to the brain of the ant (the subesophagal ganglion), where it forms a thin-walled cyst known as the brain worm. The other cercariae form thick-walled cysts. The brain worm changes the behavior of the ant, causing it to spend large amounts of time on the tips of grass blades. Here the ant is more likely to be eaten by livestock, in whose bodies parasites may continue their life cycle. This is one of many fascinating examples of parasites that manipulate the behavior of their hosts for their own benefit. For our purposes, however, the example is interesting because the brain worm, which is responsible for putting the ant in the path of a grazing animal, loses its ability to infect the mammalian host. It sacrifices its life and thereby helps to complete the life cycle of the other parasites in its group. It is hard to resist calling this kind of behavior altruistic,1 even if the parasite doesn’t think or feel anything about its fate.
How to Study Evolutionary Change
A Model of Altruistic Behavior
The question of whether and how altruism can evolve has received an enormous amount of attention from evolutionary biologists. E. O. Wilson (1975, p. 3) even called it “the central theoretical problem of sociobiology.” Evolutionists are fascinated by altruistic behaviors, not only because they might be important in nature but also because they appear so difficult to explain from the Darwinian perspective. After all, natural selection evolves traits that cause individuals to have more offspring than their competitors, not fewer. There is a selective advantage in being selfish, just as there is a selective advantage in having strong teeth and keen eyesight.
Box 1.1 presents the standard model of altruistic behavior that has been developed by evolutionary biologists. A bit of algebra is useful for precision but the basic idea can easily be described in words. Consider a population that is composed of altruists (A) and nonaltruists (S). All individuals have a certain number of offspring (a measure of fitness) in the absence of altruism. In addition, each altruist behaves in a way that decreases its own number of offspring and increases the number of offspring of a single recipient in the population. Altruists can benefit from other altruists in their group, but they also experience the cost of their own self-sacrificial behavior. Selfish types do not experience any cost and can benefit from all altruists in the group. Thus, altruists suffer a double disadvantage; not only do they incur a direct cost from performing an altruistic behavior, but they can receive donations only from other altruists, whereas a selfish individual can receive donations from all altruists. It should be obvious that selfish types always have more offspring than altruists and will be favored by natural selection. This model therefore captures the essence of what we already said in words: Altruism is the very opposite of the survival of the fittest.
Box 1.1. A mathematical model of altruistic behavior
The fitness of an individual includes both its ability to survive and its ability to reproduce. In this model, an altruistic behavior influences reproduction only, so number of offspring serves as a measure of fitness. Consider a population containing n individuals. There are two genetically encoded traits, altruism (A) and selfishness (S), which occur in frequencies p and (1 − p), respectively. The group therefore contains np altruists and n(l − p) nonaltruists. All individuals have the same average number of offspring (X) in the absence of the altruistic behavior. Each altruist behaves in a way that causes itself to have c fewer offspring and a single other member of the group to have b more offspring. The fitnesses of altruists (WA) and nonaltruists (Ws) can then be specified by the following equations:
WA = X − c + [b(np − l)/{n − 1)] (1.1)
Ws = X + [bnp/(n − 1)] (1.2)
Each altruist experiences the cost of its altruistic behavior (−c) but also serves as a possible recipient of benefits from the (np − 1) other altruists in the group. Since the other altruists are dispensing their benefits among (n − 1) individuals (to all n members except themselves), the total expected benefit that each altruist experiences is b(np − 1)/(n − 1). Nonaltruists do not experience the cost of altruism and also serve as possible recipients from all np altruists. Thus, not only do altruists suffer a direct cost (−c) but they also serve as possible recipients from fewer altruists than do selfish individuals (np − l vs. np). It is obvious that WA is always less than Ws, so altruists will always be selected against within this population.
Suppose that the parameters in the model have the following values:
| Population size (n) | 100 |
| Frequency of altruists (p) | 0.5 |
| Baseline fitness (X) | 10 |
| Benefit to recipient (b) | 5 |
| Cost to altruist (c) | 1 |
The altruistic type increases the fitness of a single recipient in its group by b = 5 units at a cost to itself of c = 1 unit. Each altruist can receive a benefit b from the 49 other altruists in the group, while selfish types can be recipients from all 50 altruists in the group. These numbers enable us to calculate the fitnesses of altruists and nonaltruists:
Fitness of altruist: WA = 10 − 1 + 5(49)/99 = 11.47
Fitness of nonaltruist: Ws = 10 + 5(50)/99 = 12.53
Everyone’s fitness is increased by the presence of altruists in the group, but the selfish S types benefit more than the altrustic A types. From these figures we can calculate the population size n′ and the frequency of altruists p′ among the offspring.
Total number of offspring: n′ = n[pWA + (1 − p)Ws] = 1200
Frequency of altruists among offspring: p′ = npWA/n′ = 0.478
The population cannot grow to infinity, so we assume that mortality operates on all types equally, returning the population to a size of n = 100 without changing the new frequency of altruism (p′ = .478). In this fashion, the altruists decline in frequency every generation and ultimately go extinct.
In the numerical example described in Box 1.1 (from Wilson 1989), we suppose that the initial population size is 100 individuals, evenly divided between altruists and nonaltruists. All individuals have 10 offspring in the absence of altruism. Each altruist bestows an additional five offspring on a single recipient at a cost of one offspring to itself. From the equations displayed in Box 1.1, we can calculate that the average A type produces 11.47 offspring and the average S type produces 12.53 offspring. For simplicity, assume that each individual reproduces by asexual reproduction, with offspring exactly resembling their parents. A total of 1,200 offspring have been produced and the proportion of altruists among the progeny is 0.478, a decline from the parental value of 0.5. Since populations cannot grow to infinity, we also assume that mortality occurs equally among the A and S types and that the population of offspring is thereby reduced to a size of 100. At this point we expect approximately 48 A types and 52 S types to survive. If this procedure is repeated many times—as it will if natural selection operates over many generations—the A type continues to decline in frequency and ultimately becomes extinct.
Before continuing, we should point out that some assumptions of this model are unrealistic. Most species are sexual, so why do we assume asexual reproduction? The answer is that the model is simpler if we do, and it is important to keep models simple. If we include sexual reproduction and get the same basic results, then we can leave sexual reproduction out for illustrative purposes. Another assumption is a one-to-one relationship between genes and behavior. Evolutionary biologists are quick to admit that this kind of genetic determinism does not exist for most species. Behaviors are caused by complex internal mechanisms that interact with the environment. These mechanisms may be genetically influenced, but recognizing this is a far cry from assuming that individuals are genetically programmed to be altruistic or selfish. Nevertheless, evolutionists frequently assume genetic determinism for the same reason that they assume asexual reproduction; it is a simplifying assumption that they hope does not alter the basic conclusions of the model. We don’t have a gene for pulling our hand away from fire, but our complex psychological mechanisms make us behave as if we do in just those circumstances in which such a gene would evolve.
Simplifying assumptions are both the soul and the Achilles heel of mathematical models. It is absolutely essential to keep a model as simple as possible to explore a given subject, such as the evolution of altruism. It often turns out, however, that a simplifying assumption leaves something out of the model that does make a difference and needs to be included to make sense of the subject. Knowing what to include and what to leave out is a subtle skill that makes theoretical biology as much an art as a science. In later chapters we will show that the assumption of genetic determinism is less innocent than it first appears (some readers may already regard it as guilty!), insofar as it obscures important possibilities for the evolution of altruism. Nevertheless, the basic problem can be exhibited most simply by assuming genetic determinism. We therefore will play along with this simplifying assumption for a while before adding more complex and reasonable connections between genes and behavior.
Returning to our model, suppose we change the equations so that A types provide one additional offspring for themselves and two offspring for everyone else in the population.2 Despite the fact that A types in this scenario increase their own fitness, they still go extinct in the model because they increase the fitness of others even more. This conclusion is well illustrated by the fable about the little red hen, who does all the work of making bread while her companions do nothing. If we change the end of the story so that her companions succeed in their freeloading ways, then everyone will gather around the table and eat the bread together. The little red hen will get some bread, perhaps even enough to repay her efforts, but her net benefit will always be less than that of her companions, who share the benefits without paying the costs. If we put this kind of interaction into our evolutionary model, the little red hen will have fewer offspring and her kind will go extinct as surely as if she decreased her absolute fitness on behalf of others. In general, evolutionary success depends on relative fitness (Williams 1966). It doesn’t matter how many offspring you have; it only matters that you have more than anyone else.
How Altruism Can Evolve
It might seem from our model that altruism can never evolve and that evolution is a process that inherently promotes selfishness. On the contrary, it is easy to show that altruism can evolve when more than one group is present. Figure 1.1 shows the simplest example of a population that is divided into two groups. The fitnesses of the A and S types are determined by the equations in Box 1.1 and equal numbers of altruists and nonaltruists exist in the total population, as in our first numerical example. In this case, however, one group has 20 percent altruists while the other group has 80 percent altruists. Looking at each group separately, we reach the same conclusion as before—selfish types will have more offspring than altruistic types. Adding the progeny of both groups together, however, we get the opposite answer; altruistic types have more offspring than selfish types.3 This outcome is strange enough that we urge the reader to inspect the numbers (see Box 1.2). There is no magic or mysticism here; the altruists increase globally, despite decreasing in frequency within each group, because the two groups contribute different numbers of individuals to the global population.
The success of altruism in Figure 1.1 is an example of a statistical phenomenon known as Simpson’s paradox (Simpson 1951; discussed in Sober 1984, 1993b). A nonbiological example of Simpson’s paradox might help explain its counterintuitive nature. During the 1970s, the University of California at Berkeley was suspected of discriminating against women in its graduate admission policies (Cartwright 1978). The percentage of women applicants who were admitted was less than the percentage of men, and the difference was large enough that it could not be attributed to chance. The University conducted a department-by-department inquiry and found in each department that women were admitted no less often than men. Evidently, women did worse than men overall, but not in any department.
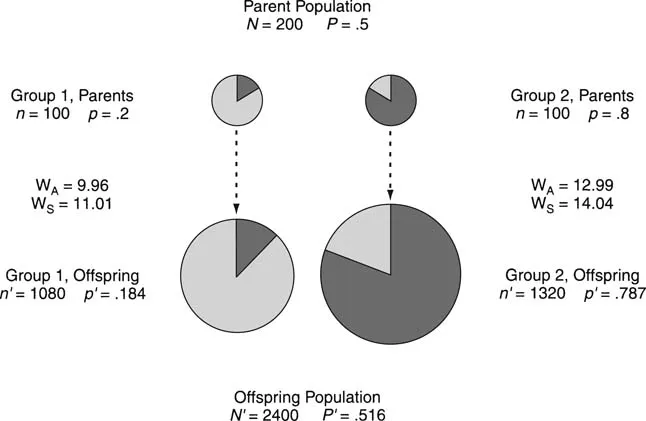
Figure 1.1. The evolution of altruistic and selfish types in an asexual population that is divided into two groups of size n = 100 (smaller circles). The altruistic type (black) makes up 20 percent of group 1 and 80 percent of group 2. In the offspring generation, the altruists decline in frequency within each group (the size of the black “slice” grows smaller), but the group with more altruists grows larger (to a population of 1,320) than the group with fewer altruists (1,080). As a result, altruists increase in frequency in the global population of 2,400 offspring. Adding the progeny from the two groups is biologically justified only if the groups periodically merge and re-form or otherwise compete in the formation of new groups.
This paradoxical finding fell into place when it was noticed that women tended to apply to departments with low acceptance rates. To see how this can happen, imagine that 90 women and 10 men apply to a department with a 30 percent acceptance rate. This department does not discriminate and therefore accepts 27 women and 3 men.
Box 1.2. The parameters of a two-group model
| Group 1 | Group 2 | |
| n | 100 | 100 |
| p | 0.2 | 0.8 |
| WA | 10 − 1 + 5(19)199 = 9.96 | 10 − 1 + 5(79)/99 = 12.99 |
| Ws | 10 + 5(20)/99 = 11.01 | 10 + 5(80)/99 = 14.04 |
| n′ | 1080 | 1320 |
| p′ | 0.184 | 0.787 |
| Global population | ||
| N | 100 + 100 = 200 | |
| P | [0.2(100) + 0.8(100)]/200 = 0.5 | |
| N′ | 1080 + 1320 = 2400 | |
| P′ | [0.184(1080) + 0.787(1320)]/2400 = 0.516 | |
Another department, with a 60 percent acceptance rate, receives applications from 10 women and 90 men. This department doesn’t discriminate either and therefore accepts 6 women and 54 men. Considering both departments together, 100 men and 100 women applied, but only 33 women were accepted, compared with 57 men. A bias exists in the two departments combined, despite the fact that it does not exist in any single department, because the departments contribute unequally to the total number of applicants who are accepted. In just the same way, altruists can increase in frequency in the two groups combined, despite the fact that they decrease in frequency within each group, because the groups contribute unequally to the total number of offspring.
Returning to the evolution of altruism, we need to emphasize that adding the progeny from the two groups is just statistical sleight of hand unless it can be justified biologically. If the two groups are permanently isolated from each other, natural selection will eliminate the altruists within each group, as we have already shown. The global increase in the frequency of altruists illustrated in Figure 1.1 will be a transient phenomenon of little interest. Suppose, however, that the progeny of both groups disperse and then physically come together before forming new groups of their own. In this case our adding procedure is appropriate and the increased frequency of altruists shown i...
Table of contents
- Cover
- Title
- Copyright
- Acknowledgments
- Contents
- Introduction: Bentham’s Corpse
- Part I. Evolutionary Altruism
- Part II. Psychological Altruism
- Conclusion: Pluralism
- Notes
- References
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Unto Others by Elliott Sober,David Sloan Wilson,Elliot Sober in PDF and/or ePUB format, as well as other popular books in Scienze biologiche & Psicologia sociale. We have over 1.5 million books available in our catalogue for you to explore.