
eBook - ePub
Cellular and Molecular Pathobiology of Cardiovascular Disease
- 338 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Cellular and Molecular Pathobiology of Cardiovascular Disease
About this book
Cellular and Molecular Pathobiology of Cardiovascular Disease focuses on the pathophysiology of common cardiovascular disease in the context of its underlying mechanisms and molecular biology. This book has been developed from the editors' experiences teaching an advanced cardiovascular pathology course for PhD trainees in the biomedical sciences, and trainees in cardiology, pathology, public health, and veterinary medicine. No other single text-reference combines clinical cardiology and cardiovascular pathology with enough molecular content for graduate students in both biomedical research and clinical departments.
The text is complemented and supported by a rich variety of photomicrographs, diagrams of molecular relationships, and tables. It is uniquely useful to a wide audience of graduate students and post-doctoral fellows in areas from pathology to physiology, genetics, pharmacology, and more, as well as medical residents in pathology, laboratory medicine, internal medicine, cardiovascular surgery, and cardiology.
- Explains how to identify cardiovascular pathologies and compare with normal physiology to aid research
- Gives concise explanations of key issues and background reading suggestions
- Covers molecular bases of diseases for better understanding of molecular events that precede or accompany the development of pathology
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MédecineSubtopic
PharmacologieChapter 1
Molecular Basis of Cardiac Development
Laura A. Dyer, PhD1, Ivan Moskowitz, MD, PhD2 and Cam Patterson, MD, MBA1, 1McAllister Heart Institute, University of North Carolina at Chapel Hill, NC, USA, 2Departments of Pediatrics and Pathology, The University of Chicago, IL, USA
Abstract
Development of the mature heart is a complex process that turns a field of mesodermal progenitors into a four-chambered organ with divided circulation. When this process goes awry, cardiac anomalies arise, and abnormalities of the heart present at birth, or congenital heart disease (CHD), have been described for centuries. CHD is highly prevalent, representing the most frequent birth defect worldwide. Tremendous strides have been made in understanding the developmental biology of cardiac morphogenesis and the human genetics of CHD. These fields of research are beginning to merge to provide an understanding of the molecular mechanisms underlying CHD. In this chapter, we present the major developmental and molecular pathways of heart formation and how they are related to human CHDs that arise when these pathways are perturbed. While CHD is generally considered conceptually as a single disease, it is much more accurate to consider CHD as a compilation of numerous distinct specific diseases, each with their own developmental and molecular etiology.
Keywords
arterial pole; atria; cardiac valves; conduction system; congenital heart defect; coronary vasculature; heart development; heart field; venous pole; ventricles
Acknowledgments
We would first like to apologize to the many authors whose studies were excluded due to space constraints. We would like to thank Dr. Andrea Portbury and Chelsea Cyr for critical reading of the manuscript.
The Heart Fields and Heart Tube Formation
The heart begins simply as a bilateral field within the lateral plate mesoderm (Figure 1.1, Table 1.1). As the early embryo undergoes formation of the gut-tube, these bilateral fields migrate toward the midline, where the cranial-most aspect of the fields will fuse ventrally to form the outer curvature of the heart tube.1 These fields will continue to migrate together, with more of the heart fields contributing to the forming tube, until the dorsal aspect of the heart fields fuse to form a closed tube.1 The initial contributors to the heart tube are known as the first heart field.2 The first heart field gives rise to the left ventricle, with some contributions to the atria and the right ventricle.3 Additional heart field progenitors from the lateral plate mesoderm continue to add to the arterial and venous poles of the heart tube; these later-adding cells are known as the second heart field. The second heart field gives rise to most of the right ventricle and atria, the most distal myocardium that surrounds the aorta and pulmonary artery, and the most proximal smooth muscle that contributes to the tunica media of the great arteries.2
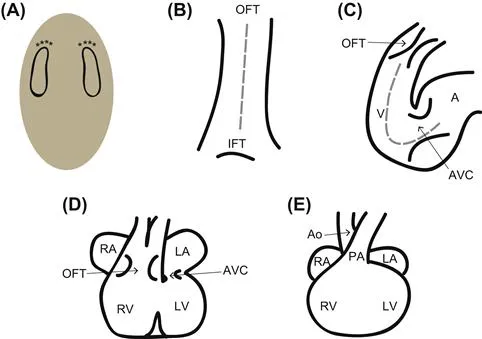
FIGURE 1.1 An overview of heart development. (A) The heart fields are specified as bilateral fields within the lateral plate mesoderm. The cranial-most aspect (asterisks) will migrate toward the midline first; this seam is depicted by the gray dashed line in B. (B) The heart tube closes ventrally, and cells continue to add from the heart fields. The dorsal aspect of the heart tube will pinch off last. After dorsal closure, additional cells can only be added via the venous (IFT, inflow tract) and arterial (OFT, outflow tract) poles. (C) As additional cells add to the heart tube, the heart tube begins to undergo looping, and the ventral midline becomes the outer curvature of the heart. During looping, the endocardial cushions begin forming in the atrioventricular canal and outflow tract. The atrioventricular canal separates the common atrium (A) from the common ventricle (V). (D) At the end of looping, the atrioventricular cushions are aligned dorsal to the outflow tract cushions, allowing connections between the left atrium and ventricle (LA and LV, respectively) and the right atrium and ventricle (RA and RV, respectively). As the outflow tract is septated, it also undergoes rotation to align the aorta with the left ventricle. Septa form between the ventricles and between the atria. (E) In the mature four-chambered heart, the aorta (Ao) serves as the outlet for the left side of the heart, and the pulmonary artery (PA) serves as the outlet for the right side of the heart.
TABLE 1.1
Major Developmental Time Points in Humans and Common Experimental Models
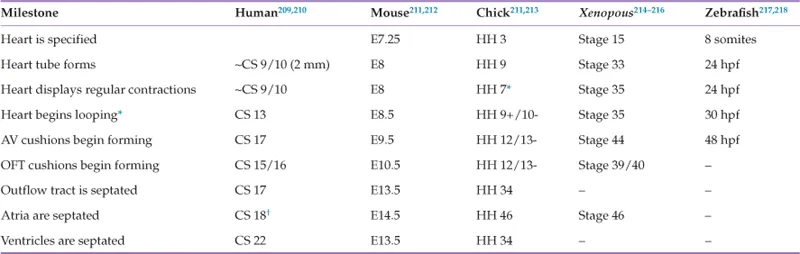
The major stages in cardiovascular development are presented for humans and the most commonly used animal models.
∗Because the heart tube begins forming as a trough that then closes dorsally, contractions are observed prior to the pinching off of the fully formed heart tube.
†The foramen ovale is still open at this stage. This fenestration is closed at the stages listed for the other species. CS, Carnegie stage219; E, embryonic day; HH, Hamburger-Hamilton stage220; Stage, Nieuwkoop and Faber stage221; hpf, hours post-fertilization. See cited references for more details.
Signaling Pathways in Heart Field Specification
The Wnt Pathway
The Wnt family includes the canonical pathway, the non-canonical pathway (also known as the planar cell polarity pathway), and the Wnt/calcium pathway (Figure 1.2). Both the canonical and non-canonical pathways have well-established roles in heart field specification. Temporal waves of canonical and non-canonical Wnt signaling play distinct roles during cardiac specification and morphogenesis. As the heart field forms from the primitive streak, Wnt3a is expressed in the primitive streak and serves as a repulsive cue to the forming heart field.4 Experiments performed in Xenopus, due to its ease of manipulation and genetic tractability,5 have shown that the early repression of Wnt signaling in the Xenopus animal cap (i.e., in the ectodermal roof of the blastocele prior to heart field specification) via Dkk-1 and Crescent is necessary for initiation of transcription of cardiac transcription factors Nkx2.5 and Tbx5 and myocardial-specific proteins troponin I and myosin heavy chain α.6 However, later in cardiac development, canonical Wnt signaling in embryonic mice at E8.75 promotes Nkx2.5, Islet1, and Baf60c within the entire heart.7 Due to the genetic similarity between Xenopus and mouse, these differences likely reflect different temporal requirements for Wnt signaling as opposed to species-specific differences.5 In the second heart field, non-canonical Wnts 5a and 11, which act through the non-canonical planar cell polarity pathway, are expressed slightly later in development and co-operatively repress the canonical Wnt pathway while also promoting expression of Islet1 and Hand2, whose expression serves to ‘mark’ the heart field;8 as such, these genes are commonly referred to as cardiac markers. Both the repression of the canonical Wnts and the induction of the heart field markers require β-catenin in the second heart field.8 Wnt5a and Wnt11 also promote proliferation within this progenitor region.8 After the heart tube forms, Wnt-stabilized β-catenin is necessary in the Islet1-expressing second heart field cells to maintain their progenitor status.9 Loss of either β-catenin or Wnt signaling in the second heart field leads to second heart field defects, including right ventricular and outflow tract defects.9,10 Even if Wnt signaling is lost under cells expressing one of the first markers of differentiated cardiomyocytes, Mesp1, second heart field proliferation is decreased, and Islet-1 expression is down-regulated.11 Conversely, overexpressing β-catenin under the Mesp1 promoter expands the Islet-1-positive second heart field and promotes proliferation.11 Later, Wnt5a specifically acts upstream of the disheveled/planar cell polarity pathway to regulate the addition of the second heart field to the arterial pole.12 In addition, Wnt signaling also promotes bone morphogenetic protein (BMP) 4 and the non-canonical Wnt 11, which promote myocardial differentiation.9,11 Thus, the Wnt pathway is critical for inducing heart field formation, maintaining progenitor status and promoting myocardial differentiation.
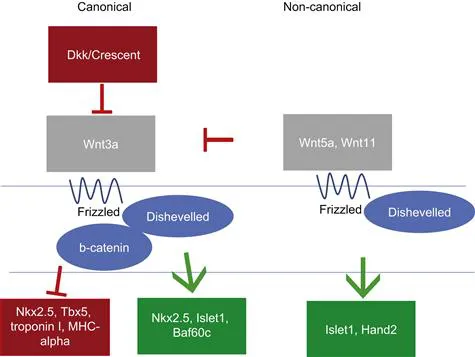
FIGURE 1.2 Wnt signaling pathway. In both the canonical and non-canonical (planar cell polarity) pathways, extracellular Wnt ligands bind to the transmembrane receptor Frizzled. In the canonical pathway, Frizzled forms a complex with Dishevelled and additional components, leading to β-catenin stabilization and translocation to the nucleus; β-catenin then promotes Wnt-induced gene expression, such as Nkx2.5 and Islet1. In contrast, in the non-canonical pathway, Frizzled and Dishevelled act on a different set of intracellular signaling molecules (e.g., Rho, Rac) to promote a different set of genes and to inhibit the canonical pathway.
Retinoic Acid
One of the earliest required signaling pathways is the retinoic acid pathway. RALDH2, the enzyme that synthesizes retinoic acid, is restricted within the lateral plate mesoderm to a region nearest to the heart field.13,14 Expression of RALDH2 progresses in a cranial–caudal direction during heart field induction and heart tube formation and establishes the posterior boundary of the heart field.15 Within the lateral plate mesoderm, retinoic acid plays an inhibitory role, where it acts both directly and indirectly to restrict cardiac transcription factors Nkx2.5 and FoxF1 to the anterior lateral plate mesoderm, Hand1 to the anterior and middle of the lateral plate mesoderm, and Sal1 to the posterior lateral plate mesoderm.13,16,17 Retinoic acid further represses GATAs 4, 5, and 6.18 This inhibitory role is required to limit the size of the heart field, and fate-mapping studies in the zebrafish, another experimental model that is valued for its genetic similarity to the mouse and ease of studying embryonic development,19 have demonstrated that zebrafish embryos with decreased levels of retinoic acid exhibit an increased number of Nkx2.5-positive cells.17 Conversely, exposing either zebrafish or Xenopus embryos to increasing levels of retinoic acid specifically leads to a reduced number of cardiomyocytes.16,17 In chick embryos, which physically develop more similar to humans as compared with mice but lack the ability to manipulate the genome as in mice,20 antagonizing retinoic acid signaling promotes the ventricular myocardial fate at the expense of the atria.15 Together, these studies suggest that retinoic acid plays two major roles in the early heart field. First, retinoic acid generally restricts the expression of heart field markers to limit the size of the heart field. Then, it specifically promotes a ‘post...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Dedication
- Foreword
- Preface
- Contributors
- Acknowledgments
- Chapter 1. Molecular Basis of Cardiac Development
- Chapter 2. Cardiac Metabolism in Health and Disease
- Chapter 3. Cardiac Atrophy and Remodeling
- Chapter 4. The Pathophysiology of Cardiac Hypertrophy and Heart Failure
- Chapter 5. Ischemic Heart Disease and its Consequences
- Chapter 6. Pathophysiology of Cardiomyopathies
- Chapter 7. Cellular and Molecular Pathobiology of the Cardiac Conduction System
- Chapter 8. Molecular Pathobiology of Myocarditis
- Chapter 9. Calcific and Degenerative Heart Valve Disease
- Chapter 10. Vasculogenesis and Angiogenesis
- Chapter 11. Diseases of Medium-Sized and Small Vessels
- Chapter 12. Pathophysiology of Atherosclerosis
- Chapter 13. Genetic Diseases of the Aorta (Including Aneurysms)
- Chapter 14. Blood Pressure Regulation and Pathology
- Chapter 15. Venous and Arterial Thrombosis
- Chapter 16. The Pericardium and its Diseases
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Cellular and Molecular Pathobiology of Cardiovascular Disease by Monte Willis,Jonathon W. Homeister,James R. Stone in PDF and/or ePUB format, as well as other popular books in Médecine & Pharmacologie. We have over 1.5 million books available in our catalogue for you to explore.