Volume I of this treatise is devoted to osmotic and ionic regulation, the physiology of excretion, and certain aspects of fish metabolism. The endocrinology of fishes is considered in Volume II. Few fields of animal physiology have expanded more rapidly during the past decade.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
B Adenohypophysial Histophysiology and Cytophysiology
II The Pituitary Gland in Teleosts
A General Organization
B Histophysiology of the Adenohypophysis
G The Neurohypophysis in Teleosts
D Blood Supply to the Teleostean Pituitary
E Hypothalamic Control of the Adenohypophysis
III The Pituitary Gland in Primitive Bony Fishes
A Superorder Chondrostei
B Superorder Holostei
IV The Pituitary Gland in Lungfishes
V The Pituitary Gland in Elasmobranchs
VI The Pituitary Gland in Cyclostomes
Acknowledgments
References
I. Introduction
A. The Pituitary Gland in General
In all vertebrates, the pituitary gland or hypophysis consists of two parts, separable on the bases of embryology, structure, and function. These are the neurohypophysis, a downgrowth from the floor of the diencephalon, and the adenohypophysis originating as an ectodermal upgrowth (Rathke’s pouch) from the roof of the embryonic buccal cavity (Wingstrand, 1966a). The two parts meet and enclose between them a mesodermal rudiment which gives rise to their intrinsic blood vessels (Green and Maxwell, 1959; Green, 1966). Thus the gland is a composite organ, and it has many different endocrine functions.
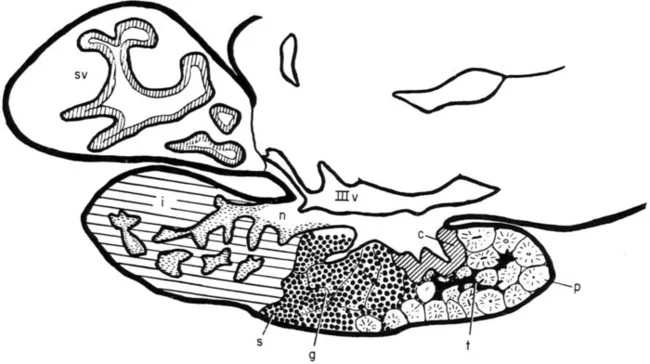
The adenohypophysis is the site of synthesis, storage, and release into the blood of several different peptide hormones; and the greater part of pituitary histophysiology is concerned with the allocation of each of these hormones to the type of pituitary cell that secretes it. The adenohypophysis is divided into the pars distalis, site of secretion of most adenohypophysial hormones, and the pars intermedia. The neurohypophysis in fishes is rather simpler than in land vertebrates (Wingstrand, 1966a) and consists essentially of a hypophysial stalk, suspending the gland from the ventral region of the diencephalon (hypothalamus) and containing an extension of the third ventricle (infundibular recess), and at the distal end of the stalk an enlargement, the neurohypophysial lobe or core, which forms the middle of the gland (Figs. 1 and 2). The stalk contains the axonal fibers of neurosecretory cells, the cell bodies being located in the hypothalamus. The neurohypophysial core consists largely of the endings of these fibers interspersed with cells termed “pituicytes.” The neurohypophysis seems to be in general a storage-release center for materials which are actually synthesized in the hypothalamus and then transported to the neurohypophysial core along the neurosecretory axons (Section II, C). In many fishes, the neurohypophysial stalk is virtually absent, the pituitary then being pressed close to the ventral surface of the hypothalamus, while in a few teleosts (e.g., Lophius) the neurohypophysial stalk is extremely long (Wingstrand, 1966a).
Fig. 1Anguilla anguilla. Diagram of midsagittal section through the eel pituitary, anterior to the right. Follicles of prolactin cells (p), mixed with TSH cells (t), and bordered posteriorly by ACTH cells (c) form the rostral pars distalis. The proximal pars distalis comprises cords of cells below the neurohypophysis (n), mainly composed of growth hormone cells or somatotrops (s) in the sexually immature fish, with scattered immature gonadotrops (g). Posteriorly is the pars intermedia (i), deeply invaded by processes of the neurohypophysis, which in this region displays masses of AF + ve neurosecretory material (horizontal shading). The saccus vasculosus (sv) projects behind the pituitary (Section II,
); IIIv indicates third ventricle, the floor and sides being the hypothalamus, the floor being also termed the “infundibular floor.”. From Olivereau (1967a)
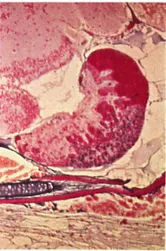
Fig. 2Poecilia latipinna. Sagittal section through pituitary of a female fish, anterior to the right. The mass of prolactin cells (bright red) together with the bands of colorless ACTH cells projecting into the neurohypophysis posteriorly form the rostral pars distalis. In the proximal pars distalis, note the ventral mass of blue gonadotrops and the dorsally projecting fingers of orange growth hormone cells, mixed with a few blue TSH cells. The pars intermedia is small in cyprinodonts and forms a thin ventral rim to the posterior neurohypophysis, which contains large Herring bodies (neurosecretory material). Azan.
B. Adenohypophysial Histophysiology and Cytophysiology
Purves (1966) has usefully divided the cytological criteria used in the study of the adenohypophysis into two groups. The first category consists of features that are indicators of the specific nature of the functions of individual cell types such as granule size, staining reactions and chemical nature, cell morphology, and reactions to specific physiological alterations; these features are the data of special cytology, which is particularly concerned with allocation of function to each cell type. The second category includes those features which are indicators of the functional state of the cell, indicating high or low rates of metabolic or secretory activity such as nuclear size, nucleolar size, amount of cytoplasmic RNA, state of the Golgi apparatus, and accumulation or loss of secretory granules. These features constitute the field of general cytology. In the study of fishes, as in other vertebrate groups, workers on the pituitary have been concerned with both kinds of criteria. However, more than in the highly worked field of mammalian pituitary histophysiology, most investigations on fishes are still primarily concerned with special cytology. This implies that there is, as yet, no complete general agreement about the functions of the various types of cell distinguishable in the fish pituitary; although perhaps with the greater technical standardizations that have come about in recent years, especially the use of methods developed in mammalian studies by Herlant (1956, 1960), and first applied to the fish pituitary by Olivereau and Herlant (1954, 1960), the actual morphological and tinctorial characteristics of the teleostean adenohypophysial cell types would now be agreed upon by most workers. However, there is still disagreement about the functions of these cell types since the experimental allocation of function to the cells has been attempted systematically in only a few species. It follows that not all workers in this field would agree on a generally applicable functional nomenclature of cell types. In this review, the nomenclature used is the one established partly on functional and partly on tinctorial grounds following experimental studies in the eel, Anguilla, and the molly, Poecilia (Olivereau, 1963b; Olivereau and Ball, 1964). It is a mixed nomenclature, in Purves’ terms (1966), based partly on similarities in staining properties of the secretory granules to those in mammalian cells of demonstrated function but mainly (primarily) on the characteristic responses of the cells in fishes to experimental situations designed to alter the secretion rates of the different adenohypophysial hormones. Logically, one is on equivocal grou...
Table of contents
Cover image
Title page
Table of Contents
Contributors
Copyright page
List of Contributors
Preface
Contents of Other Volumes
Chapter 1: The Pituitary Gland: Anatomy and Histophysiology
Chapter 2: The Neurohypophysis
Chapter 3: Prolactin (Fish Prolactin or Paralactin) and Growth Hormone
Chapter 4: Thyroid Function and Its Control in Fishes
Chapter 5: The Endocrine Pancreas
Chapter 6: The Adrenocortical Steroids, Adrenocorticotropin and the Corpuscles of Stannius
Chapter 7: The Ultimobranchial Glands and Calcium Regulation
Chapter 8: Urophysis and Caudal Neurosecretory Systemxs
Author Index
Systematic Index
Subject Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
Both plans are available with monthly, semester, or annual billing cycles.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go. Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access The Endocrine System by in PDF and/or ePUB format, as well as other popular books in Physical Sciences & Geophysics. We have over 1.5 million books available in our catalogue for you to explore.