
eBook - ePub
Atlas of Early Zebrafish Brain Development
A Tool for Molecular Neurogenetics
- 258 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Atlas of Early Zebrafish Brain Development
A Tool for Molecular Neurogenetics
About this book
Atlas of Early Zebrafish Brain Development: A Tool for Molecular Neurogenetics, Second Edition, remains the only neuroanatomical expression atlas of important genetic and immunohistochemical markers of this vertebrate model system.
It represents a key reference and interpretation matrix for analyzing expression domains of genes involved in Zebrafish brain development and neurogenesis, and serves as a continuing milestone in this research area.
This updated volume provides in-situ hybridized and immunostained preparations of complete series of brain sections, revealing markers of the fundamental stages in the life history of neuronal cells in very high quality preparations and photographic plates. Specific additions to this edition include documentation on the distribution of neurons expressing GABA, dopamine and serotonin, material on the basal ganglia, hypothalamus, and the caudal, segmented part of the diencephalon, new theories on the early organization of the telencephalon and thalamus, and integration of a comparative perspective on the mid- and hindbrain.
- Documentation on the distribution of neurons expressing GABA, dopamine and serotonin
- Material on the basal ganglia, hypothalamus, and the caudal, segmented part of the diencephalon
- New theories about the early organization of the telencephalon and thalamus
- Integration of a comparative perspective on the mid- and hindbrain
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Chapter 1
Vertebrate Central Nervous System Development
Introduction
Abstract
The first chapter introduces basics of nervous system development. We start with the vertebrate-characteristic morphogenetic event of neurulation. This process leads to the internalized neural tube that forms the entire central nervous system (CNS). In parallel, the formation of neural crest and ectodermal thickenings called placodes develop into the peripheral nervous system and the majority of head sense organs. The neural tube comprises the proliferative neuroepithelium that exclusively produces all neural cell types of brain and spinal cord (including neurons, astro-, and oligodendroglia). Subsequently, neurogenetic processes such as neural proliferation, determination, and differentiation take place within the neural tube wall. In general, these neurogenetic processes include radial migration of neuronal precursors from periventricular to subpial sites. The theoretical framework of the prosomeric model explains emerging neuroanatomical differences along dorsoventral and anteroposterior central nervous axes. Furthermore, the chapter highlights the specific development of the mammalian isocortex, cerebellum, and thalamus with a focus on inside-out versus outside-in patterns of cortex formation/tangential migration events. Finally, it addresses primary (embryonic) and secondary (larval) zebrafish neurogenesis
Keywords
Determination
Differentiation
Neurogenesis
Neurulation
Proliferation
Prosomeric model
1.1 Introductory Remarks: The Order of the Universe
The individual universe each human being mentally carries around is a product of our brain, which most likely represents the most complex object in the world. How does the brain come into existence during development? The ontogeny of the vertebrate central nervous system (CNS: brain and spinal cord) itself is a highly complex yet strongly conserved morphogenetic process. The early vertebrate embryo exhibits three embryonic cellular sheets called germ layers, the most peripheral one being the ectoderm. Its dorsal portion represents the neuroectoderm that develops into the prospective nervous system. The neuroectoderm segregates early from the ventrolaterally lying general ectoderm, which forms the future epidermal skin and its derivatives. Subsequently, the central main portion of the neuroectoderm, the neural plate, is “swallowed” by the general ectoderm through the process of neurulation. As a result, the neural plate separates from the general ectoderm and descends into the deep of the embryo, where it develops into a hollow neural tube surrounding a cerebrospinal fluid- (liquor-) filled ventricle. Meanwhile, the general ectoderm closes the temporarily opened dorsal backside of the embryo. The apparent formation along the anteroposterior axis of initially three brain vesicles (forebrain, midbrain, hindbrain) that transform subsequently into five brain vesicles (telencephalon, diencephalon, mesencephalon, metencephalon, myelencephalon) somewhat hides the parallel establishment of finer transverse subdivisions called neuromeres (rhombomeres in the hindbrain, prosomeres in the forebrain, see later). What follows is the differentiation of functional subdivisions of the brain and spinal cord (for example, the optic tectum/superior colliculus, a midbrain sensorimotor integration center, or the hypothalamus, the major control center of visceral processes).
These events of vertebrate CNS formation are accompanied by the emigration of the most lateral portions of the neuroectoderm, the neural folds (Neuralwülste) and some directly adjacent neuroectodermal regions, which are not incorporated into the neural tube during neurulation. These neuroectodermal components comprise the neural crest of the head and trunk and the placodes that are located in the head only. Eventually neural crest and placodes give rise to cells that migrate to their final destination in the adult body, where they form almost all sensory organs and the sensory neurons in dorsal root and cranial nerve ganglia, which innervate them, and the visceral ganglia of the autonomic nervous system. These somatic and visceral ganglia and their associated nerves represent the peripheral nervous system (PNS).
For many decades, scientists have investigated these complex developmental events leading to the formation of a nervous system in various vertebrates, especially in model animals, such as in the African clawed frog Xenopus, the chick, rodents like the mouse and rat, and most recently, the zebrafish, at molecular, cellular, histological, or morphological levels. Thus, a wealth of scientific literature is available, ready to satisfy almost any hunger for information.
Why then the present book? First of all, it integrates knowledge on CNS development coming from classical developmental studies with recent molecular data related to neuro- and gliogenesis. In particular, we will demonstrate the detailed spatiotemporal order of early postembryonic zebrafish brain development using molecular and cellular markers of neuro- and gliogenesis to visualize local differences, embedded in a holistic morphogenetic context. By combining old and new knowledge, we provide a neuroanatomically based molecular atlas of neural development in the zebrafish brain. We hope that such information—beyond delivering a refined molecular neuroanatomy—will prove to be enlightening in the future elucidation of neurogenetic pathways and their mechanisms.
1.2 Major Developmental Stages of the Vertebrate Neural Tube
Historically, scientists have always aimed to identify the fundamental building blocks of the vertebrate brain. In other words, they have tried to determine the basic developmental brain units and to answer the question of how these transform into functional structures of the mature brain. Before going into a more detailed discourse of the development of finer brain subdivisions (see Section 1.3), we will shortly examine the early events of brain morphogenesis relating to the emergence of vesicles and neuromeres, as well as of longitudinal zones, followed by an account on the neuromeric (prosomeric) model and on central nervous neurogenesis. An excellent in-depth historical account on the understanding of vertebrate brain morphogenesis has been given by Nieuwenhuys (1998a,b,c).
1.2.1 Vesicles, Neuromeres, and Longitudinal Zones
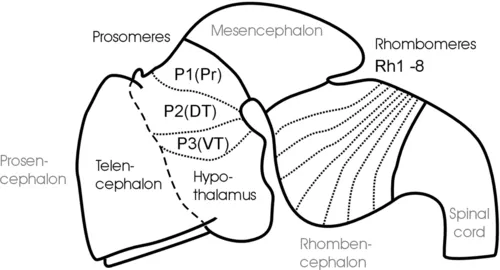
The classical description of a sequential appearance of two, three, and then five brain vesicles (i.e., transverse elements) along the anteroposterior vertebrate neural tube axis dates back to von Baer (1828). This view, however, gained popularity in the early twentieth century with the works of von Kupffer (1906) and Johnston (1909). The two-vesicle stage comprises a combined forebrain (prosencephalon) and midbrain (mesencephalon) vesicle, which is set apart by a vertical neural tube constriction from the hindbrain (rhombencephalon) vesicle. This is followed by the three-vesicle stage displaying forebrain, midbrain, and hindbrain vesicles (see Fig. 1) and, finally, by the five-vesicle stage, consisting of telencephalon and diencephalon (together forming the forebrain), mesencephalon, as well as metencephalon and myelencephalon (together forming the rhombencephalon).
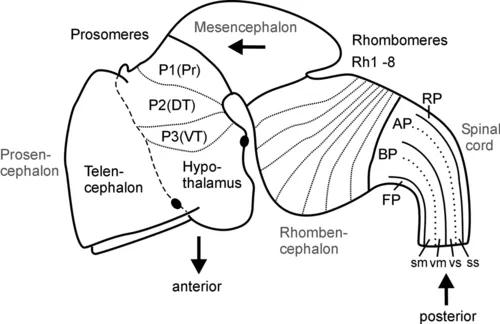
Fig. 1 Schematic lateral view of early mouse brain (E 12.5-13.5) shows prosomeric interpretation (Puelles and Rubenstein, 1993, 2003), including transverse (neuromeres) and longitudinal (columns) elements. Arrows designate anteroposterior axis of neural tube. Abbreviations: AP: alar plate; BP: basal plate; DT: dorsal thalamus (thalamus), FP: floor plate; P1-P3: prosomeres 1-3 (for more details see text); Pr: pretectum; Rh1-Rh8: rhombomeres 1-8; RP: roof plate; sm: somatomotor column; ss: somaotosensory column; T: telencephalon; vm: visceromotor column; vs: viscerosensory column; VT: ventral thalamus (prethalamus).
The subdivision of both forebrain and hindbrain into two separable (transverse) vesicles, each along the anteroposterior axis, arguably is an epiphenomenon, visible only in mammalian embryos that show an enormous early growth of the telencephalic hemispheres. This hypertrophy detracts from the fact that the most rostral vertebrate neural tube in reality includes in addition to the dorsally lying telencephalon also ventrally the smaller hypothalamus. This renders the concept of transverse telencephalic and diencephalic vesicles obsolete because part of the diencephalon (the hypothalamus) is included in a transverse unit with the telencephalon (Puelles and Rubenstein, 1993, 2003). Also, the somewhat later, characteristic mammalian emergence of a large cerebellum and ventral pontine region led to the impression that the medulla oblongata develops from two separable vesicles, the metencephalic one exhibiting cerebellum and pons, and a myelencephalic vesicle posteriorly. Indeed, the dorsal portion of the anteriormost rhombomere specializes into a dorsal cerebellum. In contrast, the ventral pontine region, which relays cortical information to the cerebellum, develops from ventral and fused portions of rhombomeres 1-4 (Aroca and Puelles, 2005; Alonso et al., 2012). This complex development of the anterior hindbrain historically resulted in the misinterpretation that the two enlargements of cerebellum and pons form a true transverse unit, which was called the metencephalon to separate it from the posterior myelencephalon. However, the medulla oblongata does not show a real boundary between these two postulated vesicles; instead, it is rather divided into a higher number of early segmental entities, the rhombomeres (see later discussion and Fig. 1).
In contrast, the three-vesicle stage reflects on fundamental anteroposterior vertebrate brain divisions. Especially the midbrain-hindbrain boundary has been strongly corroborated in modern developmental neurobiology as a singularly definable boundary and signaling center in the vertebrate brain (Marín and Puelles, 1994; Bally-Cuif and Wassef, 1995; Brand et al., 1996; Lumsden and Krumlauf, 1996; Reifers et al., 1998; Wurst and Bally-Cuif, 2001). Similarly, there is clear evidence for cellular lineage restriction (Larsen et al., 2001) and signaling center function (Scholpp and Brand, 2003) at the forebrain-midbrain boundary.
Equally important for the understanding of the vertebrate CNS bauplan are two neu...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Preface 2015
- Preface 2005
- Acknowledgments
- Chapter 1: Vertebrate Central Nervous System Development: Introduction
- Chapter 2: Atlas of Cellular Markers in Zebrafish Neurogenesis: Atlas
- Chapter 3: Interpretation of Data—How to Use the Atlas: Analysis
- Chapter 4: Comparison of Vertebrate Model Systems: Model Systems
- References
- Abbreviations
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Atlas of Early Zebrafish Brain Development by Dr. Thomas Mueller,Mario Wullimann in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Biology. We have over 1.5 million books available in our catalogue for you to explore.