
eBook - ePub
Cancer Genomics
From Bench to Personalized Medicine
- 510 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Cancer Genomics addresses how recent technological advances in genomics are shaping how we diagnose and treat cancer. Built on the historical context of cancer genetics over the past 30 years, the book provides a snapshot of the current issues and state-of-the-art technologies used in cancer genomics. Subsequent chapters highlight how these approaches have informed our understanding of hereditary cancer syndromes and the diagnosis, treatment and outcome in a variety of adult and pediatric solid tumors and hematologic malignancies. The dramatic increase in cancer genomics research and ever-increasing availability of genomic testing are not without significant ethical issues, which are addressed in the context of the return of research results and the legal considerations underlying the commercialization of genomic discoveries. Finally, the book concludes with "Future Directions", examining the next great challenges to face the field of cancer genomics, namely the contribution of non-coding RNAs to disease pathogenesis and the interaction of the human genome with the environment.
- Tools such as sidebars, key concept summaries, a glossary, and acronym and abbreviation definitions make this book highly accessible to researchers from several fields associated with cancer genomics.
- Contributions from thought leaders provide valuable historical perspective to relate the advances in the field to current technologies and literature.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Part 1
Introduction
Outline
Chapter 1 Historical Perspective and Current Challenges of Cancer Genomics
Chapter 1
Historical Perspective and Current Challenges of Cancer Genomics
Graham Dellaire1 and Robert J. Arceci2, 1Departments of Pathology and Biochemistry & Molecular Biology, Dalhousie University, Halifax, NS, Canada, 2Children’s Center for Cancer and Blood Disorders, Hematology/Oncology and The Ron Matricaria Institute of Molecular Medicine, Phoenix Children’s Hospital, Department of Child Health, University of Arizona, College of Medicine, Phoenix, AZ, USA
From the first descriptions of cancer in Egypt around 3000 BC to our current “one week” whole-genome sequence, the history of integrating new ideas into the practice of medicine has been unrelenting, although not without its failures as well its successes. This chapter represents a brief historical summary of some of the key success stories in our understanding of cancer that has led to our current age of cancer genomics. As the Chinese proverb states, “When you drink from the well, remember who dug it”, and reflecting on this rich and varied history, we conclude the chapter with a discussion of current and future challenges to the application of our new and developing understanding of cancer genomes to patient therapy.
Keywords
chromothripsis; chromoplexy; DNA microarrays; DNA sequencing; epigenetics; kataegis; massively parallel sequencing; Philadelphia chromosome; RNA-sequencing
Contents
A Historical Perspective on the Development of Cancer Genomics
Current and Future Challenges
Glossary
Abbreviations
References
Key Concepts
• Advances in nucleic acid sequencing technology have had a profound impact on the field of cancer genomics and have enabled the interrogation of the genetic basis of cancer at the single nucleotide level
• Cancer genomics has provided a detailed view of the complexity of the cancer genome, including the extraordinary ability to sustain and thrive on alterations of DNA
• Translating cancer genomics into clinically real time and actionable personalized medicine is beginning to be tested, although significant advances in the speed of providing molecular data, analysis, utilizing combination targeted agents and understanding clinical response or no response will require new generations of technology and bioinformatic tools
A Historical Perspective on the Development of Cancer Genomics
Several hundred years BC, Hippocrates is attributed with providing us with the term “carcinoma” and thus “cancer”, originating possibly from the image of finger-like extensions (veins) from a tumorous (main body of the tumor) breast lesion that shared resemblance to the shape of a crab. Around 400 years later, the Roman physician Celsus translated the Greek (karkinos) into the Latin word for crab, which led to the term “cancer” [1,2]. A relative late-comer to this nomenclature narrative, in 168 BC, Galen introduced the terms “oncos”, meaning “swelling” to describe tumors, leading to the term defining the field of oncology [3,4].
The next 2000 years witnessed several key events that helped to refine further the still ongoing main areas of cancer investigation and treatment. Maimonides in AD 1190 appears to have been the first to document surgically removing tumors [3]. The recognition of cancer clustering in distinct populations was introduced in 1713 with Razmazzini’s observation of the low cervical but high breast cancer incidence in nuns [2]. The observation that environmental and occupational exposures can be associated with increased incidence of specific cancers also became evident. In the first half of the 1800s, Recmier appears to have reignited the flare for nomenclature by writing about “metastasis” in 1829 to describe the movement of some cancers to different parts of the body [3]. Muller’s notes on the cellular origin of cancer also in 1838, and Paget’s subsequent “seed and soil” hypothesis over 50 years later in 1889, established the cognitive paradigm for the cell biological basis for cancer and the concept of microenvironmental niches [2,3]. The first half of the 20th century was ushered in by a set of remarkable observations in cancer biology that were made before the discovery of DNA. These included the theories of Rous regarding the potential viral origin of some cancers in 1910, derived from his work on avian sarcomas, and the concept of the somatic mutation theory of cancer by Boveri in 1914 that stemmed from his work on polyspermic development in invertebrates [5,6].
While not necessarily providing a deeper understanding of the mechanisms of carcinogenesis, the first half of the 20th century in many ways broke open the gates of cancer treatment. After being commissioned by the US government to understand the physiological consequences of nitrogen mustard gas used in warfare, Louis Goodman and Alfred Gilman recognized the key bone marrow toxicity of this agent and subsequently introduced its intravenous use for the treatment of lymphoid malignancies in 1946 [7]. Soon afterward in 1948, the antimetabolite aminopterin was used to treat several children with acute lymphoblastic leukemia by Farber and colleagues, a treatment built on the work of the chemist Subbarao [8]. A decade later, in 1958, Hertz, Li and colleagues reported the first cure of a metastatic tumor, namely a gestational-related choriocarcinoma, with another antimetabolite, methotrexate [9].
However, despite such encouraging forays into treating patients, few cures were achievable with surgery, radiation therapy and chemotherapy. In this regard, the extraordinary efforts of Ms Mary Lasker following her husband’s death from cancer should not go unmentioned. Through her efforts and the Citizens Committee for the Conquest of Cancer through the 1960s, they challenged government, physicians and scientists to push forth with a “War on Cancer” [10–12]. And this was in spite of a significant number of naysayers who had concluded in various publications that we knew enough to cure cancer and all that was needed was to translate the knowledge that was available at the time. Such a lack of vision was thankfully thwarted by those who propitiously concluded that only through scientific discovery and its ongoing application would improvements in cancer outcomes occur. In 1971, the US National Cancer Act was passed by Congress and then President Nixon signed it into law within 2 weeks, an astonishingly rapid accomplishment on the part of government and one that should inform current, often stalled efforts [10–13]. The consequences of the above investment, along with other efforts across the globe [14], led to an infusion of intellectual engagement and financial support for conquering cancer. The results included the establishment of clinical trial groups, comprehensive cancer centers, an explosion of new anticancer agents from the lab and from nature, and the beginning of work focused on the biological understanding of cancer. This latter work built of course on the model and profound implications of the seminal discovery of the structure of the DNA double helix by Watson and Crick in 1953 [15], a discovery that would earn them the Nobel Prize in Physiology or Medicine in 1962, an accolade they shared with colleague Maurice Wilkins. In many ways, the journey to our present age of high-throughput and genome-wide discoveries in cancer biology began with this fundamental description of the fabric of life and, as such, this discovery makes a suitable origin point from which to chronicle the key events in cancer genomics in the last 60 years (Figure 1.1).
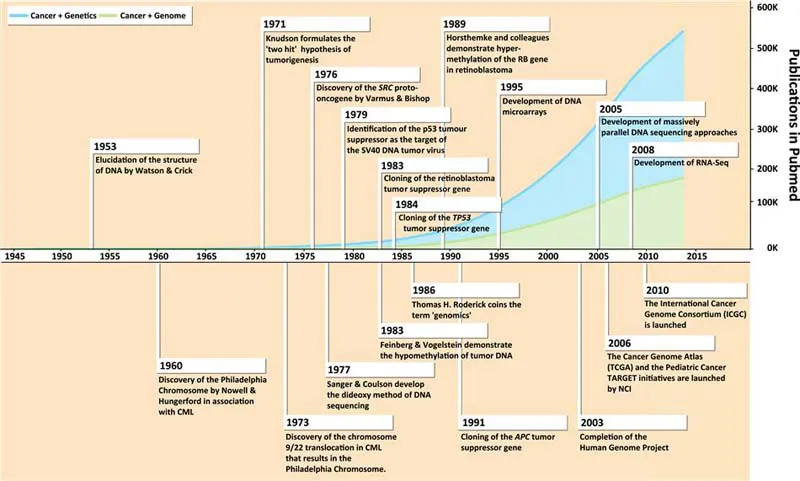
Figure 1.1 Historical milestones in cancer genomics. Key milestones in the field of cancer genomics are depicted starting with the elucidation of the structure of DNA by Watson and Crick in 1953. These milestones are depicted over a line graph of the total number of publications listed in the Pubmed database of the National Center for Biotechnology Information (NCBI) with the key-words “Cancer+(Genetics or Gene)” (in blue), or “Cancer+(Genomics or Genome)” (in green) from 1945 to 2013.
With the structure of the molecule of heredity in hand, the latter half of the 20th century saw a number of major contributions to our understanding of the biochemical and genetic underpinnings of cancer. These contributions included the identification of the “Philadelphia chromosome” as a genetic marker of chronic myelogenous leukemia (CML) by Nowell and Hungerford in 1960, and the subsequent identification of chromosomes 9 and 22 as the translocation partners underlying this anomalous chromosome by Rowley in 1973 [16,17]; the identification of the first cellular proto-oncogene, SRC, by Varmus and Bishop in the 1976 [18] leading to the realization that cellular genes could become deregulated resulting in tumorigenesis; and the identification of the p53 protein in 1979 as the primary molecular target underlying transformation by the DNA tumor virus, simian virus 40 (SV40) [18–20]. Empowered by the molecular tools developed for the study of tumor viruses, scientists studying cancer made many more seminal discoveries in the 1980s and early 1990s, including the identification of several tumor suppressor genes including retinoblastoma (RB) [21,22], the gene encoding p53 (TP53) [18–20,23,24] and adenomatous polyposis coli (APC) gene [25–27]. All these discoveries were foreshadowed by Knudson’s “two hit” hypothesis of tumorigenesis and his pioneering epidemiological studies of retinoblastoma in 1971 [28], which laid the conceptual framework for how loss of heterozygosity (LOH) of a tumor suppressor gene contributes to cancer development. It was during the 1980s that non-genetic mechanisms of oncogene regulation were first identified. One such epigenetic mechanism of gene regulation was the loss of cytosine nucleotide methylation in CpG doublets, known as hypomethylation, which was first demonstrated by Feinberg and Vogelstein in 1983 and later shown to regulate the expression of oncogenes such as HRAS [29]. Soon after, in 1986, it would be demonstrated by Baylin et al. that increased CpG...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- List of Contributors
- Part 1: Introduction
- Part 2: Genomics Technologies, Concepts and Resources
- Part 3: Hereditary Cancer Syndromes
- Part 4: Genomics of Adult and Pediatric Cancers
- Part 5: Ethics of Genomics Research
- Part 6: Future Directions
- Abbreviations
- Glossary
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Cancer Genomics by Graham Dellaire,Jason N Berman,Robert J. Arceci in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Oncology. We have over 1.5 million books available in our catalogue for you to explore.