
- 496 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Thermoregulation, Part I: From Basic Neuroscience to Clinical Neurology, Volume 154, not only reviews how body temperature regulation changes in neurological diseases, but also how this aspect affects the course and outcomes of each disease. Other sections of the volume review three therapeutic approaches that are aimed at manipulating body temperature, including induced hypothermia, induced hyperthermia and antipyretic therapy. The book is comprised of nine sections across two volumes, five dealing with the basic aspects of body temperature regulation and four dealing with the clinical aspects. Basic sections cover the Thermoregulation system, Thermoreceptors, Thermoeffectors, Neural pathways, and Thermoregulation as a homeostatic function.
In addition, the book covers the physiology and neuroanatomy of the thermoregulation system and provides descriptions of how the regulation of body temperature intervenes with other physiological functions (such as sleep, osmoregulation, and immunity), stress, exercise and aging. Basic sections serve as an introduction to the four clinical sections: Body Temperature, Clinical Significance, Abnormal Body Temperature, Thermoregulation in Neurological Disease and Therapeutic Interventions.
- Presents a clear, logical pathway from the fundamental physiology of thermoregulation, through neurobiology, to clinical applications and disease
- Enables researchers and clinicians to better understand the value of temperature measurement in disease and the use of temperature as a therapy
- Integrates content from a broad field of research, including topics on the molecular physiology of temperature receptors, to the management of accidental hypothermia
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Section III
Thermoeffectors
Chapter 9
Brown adipose tissue as a heat-producing thermoeffector
Jan Nedergaard*; Barbara Cannon Department of Molecular Biosciences, The Wenner-Gren Institute, Stockholm University, Stockholm, Sweden
* Correspondence to: Jan Nedergaard, PhD, Department of Molecular Biosciences, The Wenner-Gren Institute, Stockholm University, Stockholm 106 91, Sweden. Tel: + 46-7049-48955 email address: [email protected]
* Correspondence to: Jan Nedergaard, PhD, Department of Molecular Biosciences, The Wenner-Gren Institute, Stockholm University, Stockholm 106 91, Sweden. Tel: + 46-7049-48955 email address: [email protected]
Abstract
Extra heat for defense of body temperature can be obtained from shivering or nonshivering thermogenesis. Nonshivering thermogenesis is a facultative (i.e., only occurring when needed) and adaptive (i.e., being augmented when the demand is chronically higher) process that, in mammals, is the result of the activity of uncoupling protein-1 (UCP1) in brown and brownish adipose tissues; no other quantitatively significant mechanism that fulfills the above criteria has been established. Measurement of heat production is generally indirect, based on oxygen consumption. Heat from brown adipose tissue is generated in mammals adapted to cold, in mammalian neonates, and in mammalian hibernators during arousal; brown adipose tissue may also be active in obese mammals and thus partially protect against further obesity. UCP1 is innately inhibited by cytosolic adenosine triphosphate (ATP) and is likely activated by fatty acids released from triglycerides within the cells; this lipolysis is stimulated by norepinephrine released from the sympathetic nerves innervating the tissue. For prolonged thermogenesis, substrate is delivered by the circulation as chylomicrons, lipoproteins, fatty acids, and glucose. The proton gradient over the mitochondrial membrane created by the respiratory chain is dispersed through the activity of UCP1; brown adipose tissue is nearly devoid of ATP synthase (as compared to respiratory chain activity). UCP1 developed likely at the dawn of mammalian evolution; most mammalian species still retain functional UCP1. Other members of the uncoupling protein family cannot uncouple. Both newborn and adult humans possess active brown adipose tissue but the significance of the tissue for adult human metabolism is not established.
Keywords
UCP1;nonshivering thermogenesis;norepinephrine; nonshivering thermogenesis; norepinephrine
To defend body temperature is a prime necessity for mammalian life. Under conditions where an organism loses more heat than its basal metabolism releases as a waste product in its fight against entropy, extra heat must be produced to maintain body temperature. When such a requirement for extra heat production occurs, we – and most other mammals – have two possibilities to produce this extra heat: shivering thermogenesis and nonshivering thermogenesis. Acutely we will produce the extra heat through increased muscle tension and – given a higher demand – through shivering (see Chapter 10). However, if the demand for extra heat is chronic, most mammals will develop a capacity for nonshivering thermogenesis. It is the tenet of this chapter that all such “classic” nonshivering thermogenesis occurs in brown and brown-like adipose tissues through the functioning of the unique brown-fat protein uncoupling protein-1 (UCP1).
The basic events occurring during acute nonshivering thermogenesis are shown in Figure 9.1. As described in Chapter 17, when extra heat is needed, a nervous signal is transmitted from the brain via the sympathetic nervous system to the different depots of brown adipose tissue. When the signal reaches the tissue, release of norepinephrine occurs. This leads to breakdown of the triglycerides in the cell, as well as to an enhanced uptake of both lipids and glucose from the circulation for combustion in the mitochondria. In some way, this is also associated with the activation of UCP1, leading to heat production. The heat produced is exported from the brown adipose tissue to the rest of the organism, counteracting the heat loss.
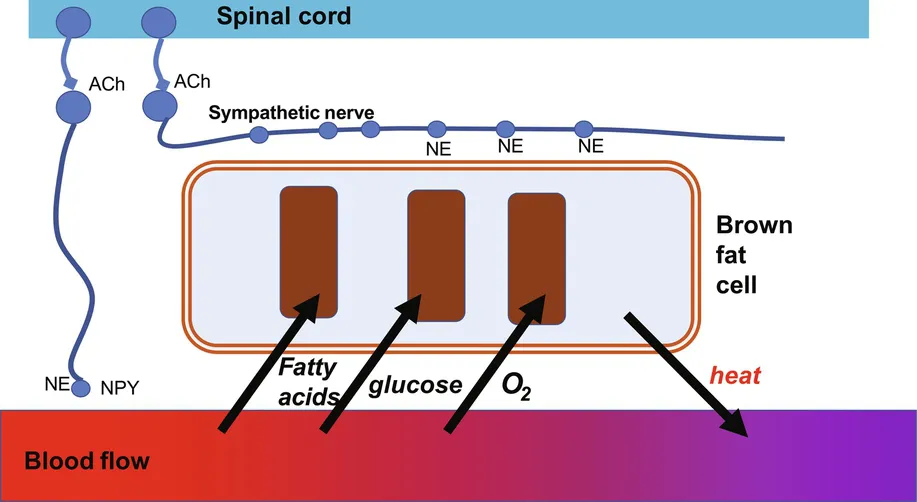
Measurement of brown-fat-derived heat production in intact animals
Direct methods
It is clearly of interest to know how much heat brown adipose tissue produces under different physiologic conditions. However, there are no simple direct ways to do this. What could theoretically be done in a given physiologic situation would be to measure the temperature difference between the incoming and the outgoing blood in the tissue and at the same time measure the volume of blood that passes through the tissue. By multiplying these parameters, the heat delivered to the body could be obtained. Even such an estimate would have some limitations: any direct transfer of heat to the surrounding tissues would be ignored, and the fact that the tissue is found in many different depots with their own blood supplies would make it almost impossible to obtain a total value for the heat produced. To date, no such direct approach has ever been accomplished.
There are other ways to approach this. One is to measure the temperature of a brown adipose tissue depot. Often the result of such a measurement is simply reported, and if the temperature increases, it is reported to indicate an activation and thus heat generation in the depot. However, the technique in itself cannot distinguish between the tissue becoming warmer due to heat produced in the tissue or becoming warmer because the entire animal is becoming warmer. The technique therefore requires monitoring of at least two temperatures in the animal, one in the brown adipose tissue and one being a reference point, such as the colon. Only if the difference between these values changes can it be established that the brown adipose tissue is heating (or is involved in heating) the animal. The limitations of the method are obviously that the data cannot be transformed into heat production, and comparisons are difficult, in that with a more recruited brown adipose tissue, the blood flow is also higher, which means that more heat is transferred away from the depot, and the temperature difference between the two sites may therefore be lower, even when heat production is actually higher.
In extension of this, the possibility of estimating brown adipose tissue temperature by following the overlying skin temperatures with thermosensitive cameras has gained popularity. In humans, this is technically feasible but in furred animals it is necessary first to shave the area that is followed, which may in itself affect the outcome. However, the main issue concerning these techniques is whether what is followed is a true reflection of heat production from depots physically underlying the skin or whether it rather reflects alterations in blood flow to the surface of the skin. There is no simple solution to this, and data obtained in this way may at best be said to be indicative of brown adipose tissue heat production. It may be added that the initial demonstration of human brown adipose tissue activity through this technique (Rothwell and Stock, 1979) does show an ephedrine-induced increase in skin temperature over the areas in the neck and clavicles where it is today recognized that adult human brown adipose tissue is located (and not at the interscapular area where at that time it would have been expected to be found).
Indirect methods
Other estimations of heat production are generally based not on the heat as such, but on the fact that the heat is the result of oxidative combustion of substrates, with the oxygen being carried to the tissue by the blood. Thus, an estimate of heat production can be made by following blood ...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Handbook of Clinical Neurology 3rd Series
- Foreword
- Preface
- Contributors
- Section I: Thermoregulation system
- Section II: Thermoreceptors
- Section III: Thermoeffectors
- Section IV: Neural pathways
- Section V: Thermoregulation as a homeostatic function
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Thermoregulation Part I by in PDF and/or ePUB format, as well as other popular books in Medicine & Neurology. We have over 1.5 million books available in our catalogue for you to explore.