
- 696 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Physiological Systems in Insects
About this book
Physiological Systems in Insects discusses the roles of molecular biology, neuroendocrinology, biochemistry, and genetics in our understanding of insects. All chapters in the new edition are updated, with major revisions to those covering swiftly evolving areas like endocrine, developmental, behavioral, and nervous systems. The new edition includes the latest details from the literature on hormone receptors, behavioral genetics, insect genomics, neural integration, and much more. Organized according to insect physiological functions, this book is fully updated with the latest and foundational research that has influenced understanding of the patterns and processes of insects and is a valuable addition to the collection of any researcher or student working with insects.
There are about 10 quintillion insects in the world divided into more than one million known species, and some scientists believe there may be more than 30 million species. As the largest living group on earth, insects can provide us with insight into adaptation, evolution, and survival. The internationally respected third edition of Marc Klowden's standard reference for entomologists and researchers and textbook for insect physiology courses provides the most comprehensive analysis of the systems that make insects important contributors to our environment.
- Third edition has been updated with new information in almost every chapter and new figures
- Includes an extensive up-to-date bibliography in each chapter
- Provides a glossary of common entomological and physiological terms
Information
Chapter 1
Signaling Systems
It is essential for cells to know where they are and what the environment around them is like. Single-celled organisms may have to detect changes in nutrients, temperature, mechanical pressure, electromagnetic fields, light, and the metabolic products of other cells of the same or different species, and make appropriate responses. This process becomes more critical for the cells of multicellular organisms, because beginning a few moments after the fertilized egg cell first divides, the individual cells must begin communicating with each other to coordinate their activities. As development proceeds, cells not only need to respond to changing conditions but also to exchange information that will determine their specialized identity and the positions they will assume in the mature organism. The information from the outside of the cell must be translated and converted to specific cellular responses in a series of steps referred to as signal transduction pathways.
In multicellular organisms these signals are most frequently chemical in nature, emanating from a signaling cell and detected by specific receptor proteins on a target cell. There are hundreds of different signal molecules, but each cell responds selectively depending on its particular function within the organism. Its response, or failure to respond, depends on whether or not it possesses a receptor for that signal. The receptor molecules are virtually always proteins that span the cell membrane. If the correct receptor receives this extracellular signal, the target cell converts it to an intracellular signal that affects cell physiology, such as changes in cell shape, metabolism, or gene expression. The external signal is thus internalized, amplified, and distributed to several internal targets (Figure 1.1). The chemical signals may consist of many types of molecules, categorized by their range and speed of activity. After the signal is transduced and arrives inside the cell, its target might be either already-synthesized proteins that become activated by the message, or changes in gene expression, either of which can alter cell physiology and behavior (Figure 1.2).
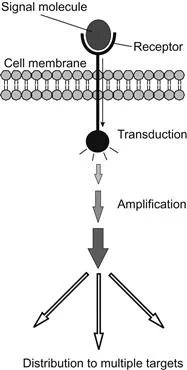
Figure 1.1 A signal molecule binds to specific receptor sites on the cell membrane. The activation of the receptor transduces the signal to the cell interior where it is amplified by parallel pathways and distributed to targets.
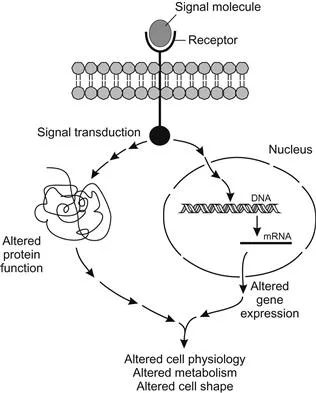
Figure 1.2 Targets of signal transduction in the cell. Cell physiology, metabolism, or behavior may be modified by altered gene expression or altered protein function.
Autocrine signaling is the most private of the signaling modes, with a cell signaling itself by producing a chemical that activates receptors within its own cytoplasm or on its surface (Figure 1.3A). An example is the prothoracic gland, which during some developmental stages activates its own production of ecdysteroids.
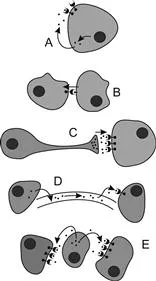
Figure 1.3 Types of cell signaling. A. Autocrine signaling. B. Contact-dependent signaling. C. Neuronal signaling. D. Endocrine signaling. E. Paracrine signaling. See the text for an explanation.
Contact-dependent signaling relies on direct contact between neighboring cells, with signaling molecules embedded in the cell membranes or passed directly through pores (Figure 1.3B). The signaling cell may produce a molecule that binds to a receptor in the adjacent target cell, or release ions that are transferred through gap junctions to trigger a response only in those cells that are in direct contact. Also called juxtacrine signaling, this is the fastest mode of communication and can be found in cardiac muscle cells whose contractions are coordinated, allowing these to occur simultaneously. Contact-dependent signaling also occurs during early development, giving adjacent cells information about their location relative to other cells, and can specify their developmental fate.
Neuronal signaling delivers messages across long distances within the organism to specific cells. As described in Chapter 11, a signaling neuron sends an electrical signal along its axon that triggers the release of a neurotransmitter at its synapse with a target cell (Figure 1.3C). The neurotransmitter binds to postsynaptic receptors in target cells, causing a physiological response in those cells. The speed of transmission is rapid, but depends on the physical distance over which the signal must travel.
Paracrine and endocrine signaling both involve the diffusion of signal molecules through an extracellular medium. Endocrine signaling is the most public, releasing hormones into the blood that are distributed to all cells of the body (Figure 1.3D). Only those cells with receptors that recognize the hormone are capable of responding. The signaling cells may consist of endocrine cells or more specialized neurosecretory cells. Some endocrine signaling molecules are hydrophobic and are able to cross the cell membrane and bind to internal receptor proteins, but most others are peptides that must remain outside and bind to external receptors.
Paracrine signaling is more intimate, with the signal molecules diffusing not through the blood but through the extracellular matrix instead (Figure 1.3E). The proteins secreted by the target cells are effective over only a short distance and induce changes only in neighboring cells that bear specific receptors localized in regions of cytoplasmic extensions called cytonemes. Because many of the signal molecules involved are proteins that are unable to enter cells through the plasma membrane, pathways of signal transduction operate within the target cells for their activation, with an extracellular receptor communicating the signal to the cell interior.
The remainder of this chapter is devoted to several important paracrine and endocrine systems in insects.
Paracrine Signaling
There is a remarkable conservation of protein messengers used for many diverse processes throughout the animal kingdom. Very similar factors operate in both mammals and insects, and, with an even more beautiful parsimony, are used at different times for different developmental events within the same organism. These are the morphogens, which diffuse short distances over fields of cells and regulate gene transcription. They have been grouped into four protein families on the basis of their structural similarities.
Fibroblast growth factors (FGF) are important during embryonic development, tissue construction, and regeneration after wounding. An example is the Breathless FGF protein that activates the Branchless FGF receptor protein (FGFR) to regulate terminal branching in the developing tracheal system of Drosophila. An FGF protein binds to an FGFR and activates a kinase that phosphorylates proteins within the responding cell. One transduction cascade initiated by FGFs is the RTK (receptor tyrosine kinase) transduction pathway, where a specific paracrine factor ligand binds to the extracellular portion of the receptor and induces a conformational change that activates the kinase activity of the cytoplasmic portion and autophosphorylates key tyrosine residues (Figure 1.4). This then recruits proteins functioning as adaptors that couple the receptor to other proteins, which can activate still other proteins to pass the message along. One of these important proteins is Ras, which exchanges its bound GDP for GTP and then stimulates subsequent molecules in the pathway, including the mitogen-activated protein kinases (MAPK), to initiate a series of phosphorylations that ultimately are transferred to the nucleus to phosphorylate certain transcription factors. The Drosophila insulin receptor is a receptor tyrosine kinase with an astonishing similarity to the human insulin receptor.
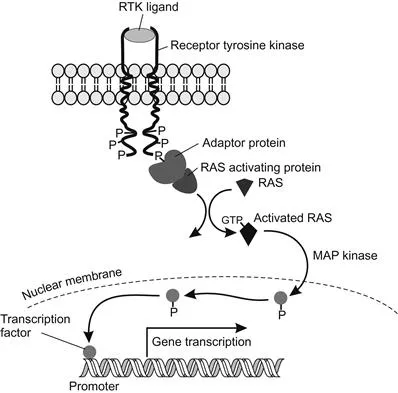
Figure 1.4 Receptor tyrosine kinase (RTK) transduction associated with a typical enzyme-linked receptor. The cytoplasmic domain of the receptor is switched on when a ligand binds to its extracellular domain. The cascade activates Ras when GTP is bound, activating a phosphorylation cascade that carries the signal to the nucleus via a MAP kinase cascade, affecting transcription factors and gene expression.
Another signal transduction cascade activated by FGFs to transmit extracellular signals to the nucleus is JAK-STAT (Janus kinase-signal transducer and activator of transcription). The cytokine receptors involved activate their associated tyrosine kinases (JAKs), which in turn activate gene regulatory proteins (STATs) that migrate to the nucleus to affect gene transcription. There are seven STAT genes in mammals, each of which binds to a different promoter, and four genes associated with the JAK family. These regulate cell proliferation, blood cell development, and the immune response, and are central to the proper growth and development of mammalian tissues. The cascade is responsible for the differentiation and renewal of both mammalian and Drosophila stem cells. In mammals, its disruption can cause leukemia and immunological disorders. The first identification of JAK-STAT signaling in insects was as a regulatory mechanism in embryonic segmentation, and it is now known to be involved in immune responses and hemocyte development, the formation of the eyes, wings, and legs, and in sex determination. It thus plays a major role in organ morphogenesis and cell rearrangement during development. Suppression of the JAK-STAT pathway in mosquitoes that are normally susceptible to dengue virus infection makes them even more susceptible to virus infection.
At least three ligands have been identified in Drosophila: Unpaired (UDP), UDP2, and UDP3. These bind and induce a conformational change in the transmembrane receptor Domeless (DOME), which phosphorylates and activates the JAK kinase Hopscotch (HOP) and transfers the phosphate group to a tyrosine residue on the transcription factor, dimerizing the STAT and allowing it to enter the nucleus where it binds to particular regions of DNA (Figure 1.5).
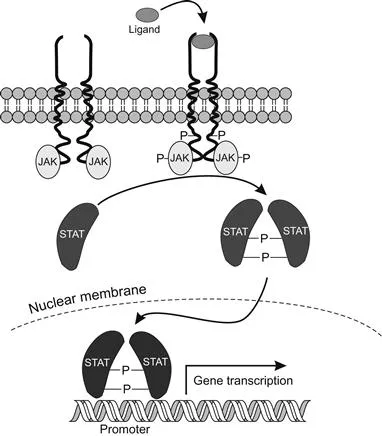
Figure 1.5 The JAK-STAT transduction cascade. The activation of JAK by the cytokine receptor subsequently activates the STAT that ...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Dedication
- Preface to the Third Edition
- Chapter 1. Signaling Systems
- Chapter 2. Integumentary Systems
- Chapter 3. Developmental Systems
- Chapter 4. Reproductive Systems
- Chapter 5. Behavioral Systems
- Chapter 6. Metabolic Systems
- Chapter 7. Circulatory Systems
- Chapter 8. Excretory Systems
- Chapter 9. Respiratory Systems
- Chapter 10. Locomotor Systems
- Chapter 11. Nervous Systems
- Chapter 12. Communication Systems
- Glossary
- Index
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Physiological Systems in Insects by Marc J. Klowden,Marc J Klowden in PDF and/or ePUB format, as well as other popular books in Biological Sciences & Medical Microbiology & Parasitology. We have over 1.5 million books available in our catalogue for you to explore.