
eBook - ePub
Microbial Energy Conversion
The Proceedings of a Seminar Sponsored by the UN Institute for Training and Research (UNITAR) and the Ministry for Research and Technology of the Federal Republic of Germany Held in Göttingen, October 1976
- 646 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Microbial Energy Conversion
The Proceedings of a Seminar Sponsored by the UN Institute for Training and Research (UNITAR) and the Ministry for Research and Technology of the Federal Republic of Germany Held in Göttingen, October 1976
About this book
Microbial Energy Conversion documents the proceedings of a seminar in Gottingen in October 1976. This book discusses the potential of microorganisms to use solar energy or convert biomass produced by solar energy in such a way that new microbial energy sources can supplement or partially replace conventional sources. This compilation reviews biomass production and elaborates on in detail the microbial processes that are involved in the conversion of the primary biomass—either freshly harvested or disposed of as waste—into energy sources that are similar to hydrogen, methane, propane, gasoline, Diesel oil, methanol, ethanol, or electricity. The microbial processes that contribute to the development of known energy resources, such as mining of low grade ores of copper, zinc, and uranium; reclamation of oil from oil shale; and recovery of conventional and heavy oil and gas, are also deliberated. This text likewise elaborates on the study of photosynthetic enzyme systems, hydrogenase, immobilization of enzymes and pigments on membranes, and construction of artificial photosynthetic units. This book is beneficial to students and researchers conducting work on microbial energy conversion.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Methane formation and cellulose digestion
ACETATE, A KEY INTERMEDIATE IN METHANOGENESIS
R.A. Mah, Center for the Health Sciences, University of California, Los Angeles, California 90024, USA
R.E. Hungate and Kyoko Ohwaki, Department of Bacteriology, University of California, Davis, California 95616, USA
Abstract
Acetate is the chief precursor of methane in anaerobic fermentations. Methane can be formed from H2 and CO2 by most methanogenic bacteria, but these substrates can also be converted to acetate by Clostridium aceticum, isolated from sludge. Its activity may increase the fraction of methane arising from acetate. Anaerobic 2% Ca acetate mineral enrichments forming methane were transferred weekly with a one-sixth volume inoculum. The rate of methane production was 9 μmoles ml−1 day−1. Methanosarcina barkeri was the chief methanogen in the enrichment but numerous non-methanogens were present, three of which were isolated. In pure cultures of M. barkeri in medium supplemented with trypticase and yeast extract the rate of methanogenesis was 20 ymoles ml−1 day−1, and similar supplementation of the enrichment culture increased the rate from 9 to 23 μmoles ml−1 day−1. The rate of methane production was affected not only by M. barkeri but also by the non-methanogens in the enrichment. Microbiological analyses of methanogenic ecosystems is recommended as a means to provide clues of value in practical applications.
It is a pleasure to participate in a symposium exploring possibilities to augment world energy sources by exploiting microbial activities. We are greatly indebted to Dr. Schlegel and the Institut für Mikrobiologie for organizing this meeting and appreciate the financial support of the United Nations Institute for Training and Research and the Bundesministerium für Forschung und Technologie.
The preceding papers have reviewed the photosynthetic storage of renewable light energy in chemical form by various ecosystems. An inventory of energy resources shows (table 1) that sunlight constitutes more than 99.94% of the total energy (excluding nuclear) available annually on Earth.
Table 1
Potential Non-nuclear World Power
| Kind | Amount in Kcal year−1 |
| Geothermal | 0.008 |
| Tidal | 0.09 |
| Water power | 20 |
| Coal, oil and gas | 80 |
| Sunlight | 170000 |
Although only a small fraction, 0.12%, of the light is used in photosynthesis the energy stored in chemical form (table 2) is about twice the annual fuel consumption.
Table 2
Maximum World Agricultural Production
| Kind | Amount in kcal year−1 |
| Arable lands | 120 |
| Grazing lands | 25 |
| Forests | 23 |
| Seas | 32 |
| Total | 200 |
This session on methanogenesis will explore potentials and problems in microbial generation of methane from available agricultural products. The processes to be considered are anaerobic. Aerobic microbial metabolism of organic matter converts energy chiefly into the form of microbial cell bodies whereas anaerobically cells are minor and the reduced carbon in the waste fermentation products contains most of the substrate energy, available through oxidation. In a complete anaerobic breakdown of organic matter carbon atoms are converted to their most reduced or most oxidized form, i.e., to the gases, CH4 or CO2.
For anaerobic reactions to occur the substrate carbon atoms must be in an intermediate oxidation state, and energywise the most favorable condition for rapid and complete conversion of the organic matter to CO2 and CH4 is provided when the C, H, and O atoms are in the ratio of CH2O, i.e. carbohydrate. The conversion of carbohydrate to CH4 and CO2 is accompanied by a ten percent decrease in the potential energy, but ninety percent of the energy available through oxidation of the initial carbohydrate can be derived through oxidation of the reduced fermentation product, methane.
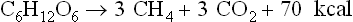
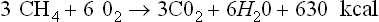
Proteins, fats and many other compounds can also be converted to CO2 and CH4 but they are not as abundant as carbohydrates, and the proteins are too valuable to be used as fuel. Lignins, comprising up to 30% of woody plant material, are a handicap to complete anaerobic conversion of plant materials. Lignins are relatively indigestible anaerobically, and their intimate association with carbohydrates reduces the digestibility of the carbohydrate components in the lignified plants. Thus carbohydrates, and possibly fats, are the chief renewable substrates suitable for anaerobic conversion to CO2 and CH4.
A carbohydrate such as hexose can be converted to methane in the overall process summarized in equation 1, but this includes two separate pathways by which methane originates. The two triose molecules derived from the hexose are fermentated in a fashion such that methane arises either by reduction of CO2 (Barker, 1936a)or by the splitting of acetate (Buswell and Sollo, 1948). Equations 3 to 5 illustrate how the hydrogen to reduce CO2 is obtained during breakdown of the two triose molecules derived from hexose.
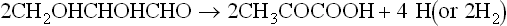
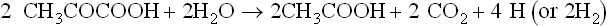
The hydrogen made available in these reactions is used to reduce CO2 to CH4.
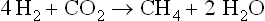
In summary,
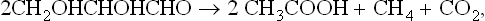
using one-third of the available hydrogen to produce one methane molecule. The acetate is then split to produce the remaining two-thirds of the methane.
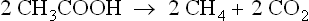
Many fermentations give rise to products such as ethanol, or propionic and butyric and higher acids, converted in sludge to methane, in part by reduction of CO2 with the hdyrogen obtained by dehydrogenation of the substrate to acetate, in part by splitting of the acetate into methane and CO2. With these fermentations also, acetate constitutes two-thirds of the carbohydrate initially attacked, and is the source of two-thirds of the methane. If a fat such as stearate is the initial substrate it can be converted by β-dehydrogenation to acetate, with the hydrogen used to reduce exogenous CO2 to methane. In this process the fraction of methane arising from acetate is slightly less than 69% of the total.
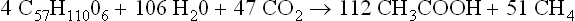
In the sludge fermentation of municipal and domestic wastes, tracer studies indicate that the fraction of methane arising from acetate is about 73% (Jeris and McCarty, 1965; Smith and Mah, 1966), 4 and 6% higher than the theoretical values for fats and carbohydrates, respectively. A possible explanation for this higher value is the occurrence in sludge of the acetigenic reaction discovered by Wieringa (1938) in which H2 and CO2 are converted into acetic acid by a spore-forming bacterium, Clostridium aceticum, isolated from Dutch canal mud.
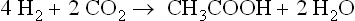
One of us (KO) has isolated presumably similar bacteria from sewage sludge (Ohwaki, 1975) at Davis. Mylroie (1953) earlier showed at Pullman that H2 and CO2 added to sewage sludge were actively converted to acetate with little formation of methane. The numbers of acetigenic bacteria at Pullman were not large, ranging from 106 to 108 ml−1. In pure culture this species grows rapidly, a ml of culture absorbing as much as 130 μmoles of H2 day−1. Yeast extract is required for growth, and vitamin B12, folate, Se, Fe and Mo are stimulatory. With this supplemented medium growth on 80% H2-20% CO2 is two to three times as great as on 80% N2-20% CO2.
Growth on the yeast extract is very rapid, and i...
Table of contents
- Cover image
- Title page
- Table of Contents
- Other Pergamon Titles of Interest
- Copyright
- PREFACE
- INTRODUCTION TO RECOMMENDATIONS
- WORKING GROUP I: Biomass
- WORKING GROUP II: Recycling of Wastes
- WORKING GROUP III: Methane Production
- WORKING GROUP IV: Photoproduction of Hydrogen/Purple Membrane
- WORKING GROUP V: Microbial Recovery of Hydrocarbons
- WORKING GROUP VI: Prices of important substrates and economics of chemical interconversions
- Primary Production of Biomass
- Methane formation and cellulose digestion
- Hydrogen formation
- Reclamation of hydrocarbons
- Engineering, operation and economics of biodigesters
- Ecological problems and waste recovery
- Ecological problems, secondary implications included
- Economic considerations
- Sociological implications in developed and developing countries
- Authors Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Microbial Energy Conversion by H. G. Schlegel,J. Barnea in PDF and/or ePUB format, as well as other popular books in Technology & Engineering & Anthropology. We have over 1.5 million books available in our catalogue for you to explore.