
- 528 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
About this book
Peripheral Neuropathies compiles lectures presented at the Symposium on Peripheral Neuropathies held in Milan, Italy on June 26-28, 1978. This book covers the basic problems in peripheral nerve pathology; axonal transport; neuropathies in chronic renal failure; peripheral neuropathies in diabetes; compression neuropathies; and metabolic, genetic, and inflammatory neuropathies. This text also discusses topics such as filamentous inclusions in Schwann cell nuclei; autonomie dysfunction and myokymia in gold neuropathy; study of the effect of gangliosides on experimental carbon disulfide neuropathy; and peripheral nerve changes in Refsum's disease before and after treatment. This compilation is a useful source of information for students and researchers conducting investigative work on peripheral neuropathy.
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicinaSubtopic
Enfermedades y alergiasAXONAL TRANSPORT
Outline
Chapter 10: NEURAL CONTROL OF GENE EXPRESSION OF SKELETAL MUSCLE FIBERS
Chapter 11: THE ROLE OF CALCIUM IN AXOPLASMIC TRANSPORT IN MAMMALIAN NERVE FIBERS
Chapter 12: AXONAL TRANSPORT AND INTRACELLULAR ORGANIZATION OF SKELETAL AND CONTRACTILE STRUCTURES IN MATURING NEURONS
Chapter 13: AXONAL TRANSPORT AND PERIPHERAL NERVE DISEASE IN MAN
Chapter 14: CHANGES IN AXONAL TRANSPORT IN VARIOUS EXPERIMENTAL NEUROPATHIES
Chapter 15: AXONAL TRANSPORT IN ACRYLAMIDE NEUROPATHY
Chapter 16: EXPERIMENTAL INVESTIGATION OF ALCOHOLIC NEUROPATHY
Chapter 18: CONDUCTANCES OF RAT SKELETAL MUSCLE FIBERS DURING RECOVERY FROM CHRONIC APPLICATION OF VINCRISTINE TO THE MOTOR NERVE
NEURAL CONTROL OF GENE EXPRESSION OF SKELETAL MUSCLE FIBERS
SALVATORE METAFORA1, ROBERTO COTRUFO2, ARMANDO FELSANI1, GIANFRANCO TAJANA3, BRUNO RUTIGLIANO4, ANTONIO DEL RIO1, PIER PAOLO DE PRISCO1, MARIA ROSARIA MONSURRO’2, MARINA MELONE2 and RICARDO MILEDI5, 1Laboratorio di Embriologia Molecolare, CNR; Arco Felice; Naples; ITALY; 2Clinica Neurologica (R), 1a Facolt à di Medicina e Chirurgia; Naples; 3Istituto di Anatomia Umana Normale; 2a Facoltà di Medicina e Chirurgia; Naples; 4Laboratorio Internazionale di Genetica e Biofisica, CNR; Naples; 5Department of Biophysics; University College; London; U.K.
ABSTRACT
Experimental results are presented in favour of the hypothesis that alpha. motoneurones control the gene expression of skeletal muscle; in fact, following denervation, we found significant changes in the complexity and molecular diversity of mRNA populations isolated from the ribosomal pellet of muscle homogenate, as compared with control muscles.
INTRODUCTION
Studies on the differentiation of slow and fast motor units and on the effects of denervation, direct and crossed reinnervation, block of nerve conduction, block of axonal flow, chronic electrical stimulations at various frequencies: on the properties of muscle fibers, have all led to the conclusion that alpha-motoneurones control some steps of differentiation and maintain the differentiated state of muscle fibers (1, 2).
Up to now, one of the main questions to be answered was how does the neural control operate at the muscular level. We have formulated the hypothesis that such control operates at the level of gene expression; an observation which suggested this hypothesis was that acetylcholine hypersensitivity of denervated muscles could be prevented by administration of Actinomycin D or Cicloheximide (1), drugs which inhibit the protein synthesis at the transcriptional and translational levels, respectively.
We have tried to give an answer to the above question by investigating the effects of denervation on the complexity and diversity of mRNA populations of a mammalian muscle. Our results indicate that, following denervation, there is a clear-cut change in the programme of muscle gene expression.
MATERIALS AND METHODS
All the procedures and the materials used can not be described here for lack of space and will be reported in the full paper to be published.
RESULTS AND DISCUSSION
Following purification, by affinity chromatography on oligo-DT cellulose, of poly (A)+mRNA’s extracted from muscle ribosomal pellets, a big incubation of control or denervated poly(A)+mRNA’s in a mRNA dependent cell free protein synthesising system from wheat germ, in the presence of 35S-methionine, was performed. (3). Following incubation, the translation products were separated on slabs of polyacrylamide-SDS gels; from the observation of the autoradiography of these gels, several differences were found between the profile obtained following incubation with poly(A)+mRNA’s from control muscles and that obtained following incubation with poly(A)+mRNA’s from denervated muscles, as shown in figure 1. This result, indicating quantitative differences in some of the sequences present in mRNA populations isolated from denervated and contralateral muscles, suggested meaningful changes in the gene expression of skeletal muscles, following denervation.
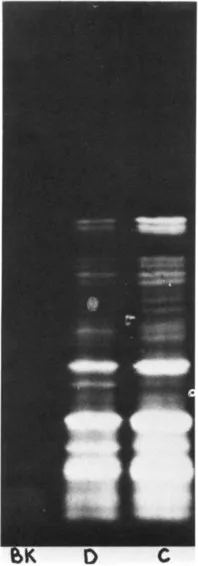
Fig. 1 Autoradiography of the electrophoretic profiles, on slabs of SDS-polyacrylamide gels, of the translation products of the incubation of muscle poly(A)+mRNA’s in a mRNA-dependent, cell free protein synthesising system, from wheat germ, in the presence of 35S-methionine. Gels contained a continuous gradient of acrylamide (9% to 15%); the run was carried out from top to bottom of the figure; from top to bottom, the bands correspond to polypeptides of decreasing molecular weight. On the left side of the slab, was carried out the electrophoresis of the blank of incubation, containing all the constituents minus mRNA’s; no incorporation of 35S-methionine was found. In the middle, were separated the translation products of poly(A)+mRNA’s from denervated muscles; on the right side, the translation products of poly(A)+mRNA’s from control muscles. In all 3 runs, 50,000 cpm were deposited on the gel. The different proportions of the translation products of poly(A)+mRNA’s from control and denervated muscles, are self evident and indicate quantitative differences of some sequences in mRNA populations.
The poly(A)+mRNA’s, isolated from control and denervated muscles, were also used as templates to synthesize, by means of a reverse transcriptase, the relevant complementary DNA (cDNA). By using the radioactive cDNA thus obtained, we have kinetically studied the molecular hybridization between homologous poly(A)+mRNA and cDNA from control and denervated muscles, as shown in figures 2 and 3. The mathematical elaboration of the kinetic curves (4,5,6), reported in tables 1 and 2 for control and denervated mRNA populations, respectively, showed that, in control mRNA populations, four different classes of abundance were present (Table 1), while, in denervated mRNA populations, only three classes of abundance were detectable (Table 2). In particular, the first class of abundance found in control mRNA populations (whose complexity was equal to 2×106 daltons), including 3 different sequences of 700,000 daltons, each repeated 3,200 times per diploid genome (see also Table 1), was not present as a single class in denervated mRNA populations (table 2); therefore the most abundant
TABLE 1
COMPLEXITY, DIVERSITY AND ABUNDANCE OF mRNA-POLY(A)+ POPULATION FROM CONTROL MUSCLE
Mathematical analysis of the hybridiz...
Mathematical analysis of the hybridiz...
Table of contents
- Cover image
- Title page
- Table of Contents
- Inside Front Cover
- Copyright
- PREFACE
- INTRODUCTORY LECTURE
- BASIC PROBLEMS IN PERIPHERAL NERVE PATHOLOGY
- AXONAL TRANSPORT
- NEUROPATHIES IN CHRONIC RENAL FAILURE
- PERIPHERAL NEUROPATHIES IN DIABETES
- COMPRESSION NEUROPATHIES
- METABOLIC, GENETIC AND INFLAMMATORY NEUROPATHIES
- MISCELLANEOUS
- AUTHOR INDEX
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Peripheral Neuropathies by N. Canal,G. Pozza in PDF and/or ePUB format, as well as other popular books in Medicina & Enfermedades y alergias. We have over 1.5 million books available in our catalogue for you to explore.