
eBook - ePub
Fundamental Neuroscience
- 1,280 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Fundamental Neuroscience
About this book
Fundamental Neuroscience, Third Edition introduces graduate and upper-level undergraduate students to the full range of contemporary neuroscience. Addressing instructor and student feedback on the previous edition, all of the chapters are rewritten to make this book more concise and student-friendly than ever before. Each chapter is once again heavily illustrated and provides clinical boxes describing experiments, disorders, and methodological approaches and concepts.Capturing the promise and excitement of this fast-moving field, Fundamental Neuroscience, 3rd Edition is the text that students will be able to reference throughout their neuroscience careers!
- 30% new material including new chapters on Dendritic Development and Spine Morphogenesis, Chemical Senses, Cerebellum, Eye Movements, Circadian Timing, Sleep and Dreaming, and Consciousness
- Additional text boxes describing key experiments, disorders, methods, and concepts
- Multiple model system coverage beyond rats, mice, and monkeys
- Extensively expanded index for easier referencing
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
II
CELLULAR AND MOLECULAR NEUROSCIENCE
CHAPTER 3 Cellular Components of Nervous Tissue
Several types of cellular elements are integrated to constitute normally functioning brain tissue. The neuron is the communicating cell, and many neuronal subtypes are connected to one another via complex circuitries, usually involving multiple synaptic connections. Neuronal physiology is supported and maintained by neuroglial cells, which have highly diverse and incompletely understood functions. These include myelination, secretion of trophic factors, maintenance of the extracellular milieu, and scavenging of molecular and cellular debris from it. Neuroglial cells also participate in the formation and maintenance of the blood–brain barrier, a multicomponent structure that is interposed between the circulatory system and the brain substance and that serves as the molecular gateway to brain tissue.
NEURONS
The neuron is a highly specialized cell type and is the essential cellular element in the CNS. All neurological processes are dependent on complex cell–cell interactions among single neurons as well as groups of related neurons. Neurons can be categorized according to their size, shape, neurochemical characteristics, location, and connectivity, which are important determinants of that particular functional role of the neuron in the brain. More importantly, neurons form circuits, and these circuits constitute the structural basis for brain function. Macrocircuits involve a population of neurons projecting from one brain region to another region, and microcircuits reflect the local cell–cell interactions within a brain region. The detailed analysis of these macro- and microcircuits is an essential step in understanding the neuronal basis of a given cortical function in the healthy and the diseased brain. Thus, these cellular characteristics allow us to appreciate the special structural and biochemical qualities of a neuron in relation to its neighbors and to place it in the context of a specific neuronal subset, circuit, or function.
Broadly speaking, therefore, there are five general categories of neurons: inhibitory neurons that make local contacts (e.g., GABAergic interneurons in the cerebral and cerebellar cortex), inhibitory neurons that make distant contacts (e.g., medium spiny neurons of the basal ganglia or Purkinje cells of the cerebellar cortex), excitatory neurons that make local contacts (e.g., spiny stellate cells of the cerebral cortex), excitatory neurons that make distant contacts (e.g., pyramidal neurons in the cerebral cortex), and neuromodulatory neurons that influence neurotransmission, often at large distances. Within these general classes, the structural variation of neurons is systematic, and careful analyses of the anatomic features of neurons have led to various categorizations and to the development of the concept of cell type. The grouping of neurons into descriptive cell types (such as chandelier, double bouquet, or bipolar cells) allows the analysis of populations of neurons and the linking of specified cellular characteristics with certain functional roles.
General Features of Neuronal Morphology
Neurons are highly polarized cells, meaning that they develop distinct subcellular domains that subserve different functions. Morphologically, in a typical neuron, three major regions can be defined: (1) the cell body (soma or perikaryon), which contains the nucleus and the major cytoplasmic organelles; (2) a variable number of dendrites, which emanate from the perikaryon and ramify over a certain volume of gray matter and which differ in size and shape, depending on the neuronal type; and (3) a single axon, which extends, in most cases, much farther from the cell body than the dendritic arbor (Fig. 3.1). Dendrites may be spiny (as in pyramidal cells) or nonspiny (as in most interneurons), whereas the axon is generally smooth and emits a variable number of branches (collaterals). In vertebrates, many axons are surrounded by an insulating myelin sheath, which facilitates rapid impulse conduction. The axon terminal region, where contacts with other cells are made, displays a wide range of morphological specializations, depending on its target area in the central or peripheral nervous system.
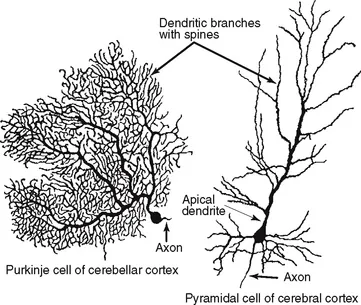
FIGURE 3.1 Typical morphology of projection neurons. (Left) A Purkinje cell of the cerebellar cortex and (right) a pyramidal neuron of the neocortex. These neurons are highly polarized. Each has an extensively branched, spiny apical dendrite, shorter basal dendrites, and a single axon emerging from the basal pole of the cell.
The cell body and dendrites are the two major domains of the cell that receive inputs, and dendrites play a critically important role in providing a massive receptive area on the neuronal surface. In addition, there is a characteristic shape for each dendritic arbor, which can be used to classify neurons into morphological types. Both the structure of the dendritic arbor and the distribution of axonal terminal ramifications confer a high level of subcellular specificity in the localization of particular synaptic contacts on a given neuron. The three-dimensional distribution of dendritic arborization is also important with respect to the type of information transferred to the neuron. A neuron with a dendritic tree restricted to a particular cortical layer may receive a very limited pool of afferents, whereas the widely expanded dendritic arborizations of a large pyramidal neuron will receive highly diversified inputs within the different cortical layers in which segments of the dendritic tree are present (Fig. 3.2) (Mountcastle, 1978). The structure of the dendritic tree is maintained by surface interactions between adhesion molecules and, intracellularly, by an array of cytoskeletal components (microtubules, neurofilaments, and associated proteins), which also take part in the movement of organelles within the dendritic cytoplasm.
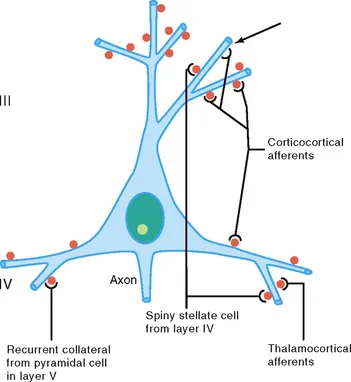
FIGURE 3.2 Schematic representation of four major excitatory inputs to pyramidal neurons. A pyramidal neuron in layer III is shown as an example. Note the preferential distribution of synaptic contacts on spines. Spines are labeled in red. Arrow shows a contact directly on the dendritic shaft.
An important specialization of the dendritic arbor of certain neurons is the presence of large numbers of dendritic spines, which are membranous protrusions. They are abundant in large pyramidal neurons and are much sparser on the dendrites of interneurons (see later).
The perikaryon contains the nucleus and a variety of cytoplasmic organelles. Stacks of rough endoplasmic reticulum are conspicuous in large neurons and, when interposed with arrays of free polyribosomes, are referred to as Nissl substance. Another feature of the perikaryal cytoplasm is the presence of a rich cytoskeleton composed primarily of neurofilaments and microtubules, discussed in detail in Chapter 4. These cytoskeletal elements are dispersed in bundles that extend from the soma into the axon and dendrites.
Whereas dendrites and the cell body can be characterized as domains of the neuron that receive afferents, the axon, at the other pole of the neuron, is responsible for transmitting neural information. This information may be primary, in the case of a sensory receptor, or processed information that has already been modified through a series of integrative steps. The morphology of the axon and its course through the nervous system are correlated with the type of information processed by the particular neuron and by its connectivity patterns with other neurons. The axon leaves the cell body from a small swelling called the axon hillock. This structure is particularly apparent in large pyramidal neurons; in other cell types, the axon sometimes emerges from one of the main dendrites. At the axon hillock, microtubules are packed into bundles that enter the axon as parallel fascicles. The axon hillock is the part of the neuron where the action potential is generated. The axon is generally unmyelinated in local circuit neurons (such as inhibitory interneurons), but it is myelinated in neurons that furnish connections between different parts of the nervous system. Axons usually have higher numbers of neurofilaments than dendrites, although this distinction can be difficult to make in small elements that contain fewer neurofilaments. In addition, the axon may show extensive, spatially constrained ramified, as in certain local circuit neurons; it may give out a large number of recurrent collaterals, as in neurons connecting different cortical regions; or it may be relatively straight in the case of projections to subcortical centers, as in cortical motor neurons that send their very long axons to the ventral horn of the spinal cord. At the interface of axon terminals with target cells are the synapses, which represent specialized zones of contact consisting of a presynaptic (axonal) element, a narrow synaptic cleft, and a post-synaptic element on a dendrite or perikaryon.
Synapses and Spines
Synapses
Each synapse is a complex of several components: (1) a presynaptic element, (2) a clef...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- Preface to the Third Edition
- About The Editors
- Contributors
- I: NEUROSCIENCE
- II: CELLULAR AND MOLECULAR NEUROSCIENCE
- III: NERVOUS SYSTEM DEVELOPMENT
- IV: SENSORY SYSTEMS
- V: MOTOR SYSTEMS
- VI: REGULATORY SYSTEMS
- VII: BEHAVIORAL AND COGNITIVE NEUROSCIENCE
- Index
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Fundamental Neuroscience by Larry Squire,Darwin Berg,Floyd E. Bloom,Sascha du Lac,Anirvan Ghosh,Nicholas C. Spitzer,Larry R. Squire,Floyd E. Bloom,Nicholas C. Spitzer in PDF and/or ePUB format, as well as other popular books in Psychology & Clinical Psychology. We have over 1.5 million books available in our catalogue for you to explore.