
- 353 pages
- English
- ePUB (mobile friendly)
- Available on iOS & Android
eBook - ePub
Antibody Techniques
About this book
The applicability of immunotechniques to a wide variety of research problems in many areas of biology and chemistry has expanded dramatically over the last two decades ever since the introduction of monoclonal antibodies and sophisticated immunosorbent techniques. Exquisitely specific antibody molecules provide means of separation, quantitative and qualitative analysis, and localization useful to anyone doing biological or biochemical research.
This practical guide to immunotechniques is especially designed to be easily understood by people with little practical experience using antibodies. It clearly presents detailed, easy-to-follow, step-by-step methods for the widely used techniques that exploit the unique properties of antibodies and will help researchers use antibodies to their maximum advantage.
- Detailed, easy-to-follow, step-by-step protocols
- Convenient, easy-to-use format
- Extensive practical information
- Essential background information
- Helpful hints
Trusted by 375,005 students
Access to over 1.5 million titles for a fair monthly price.
Study more efficiently using our study tools.
Information
Topic
MedicineSubtopic
Immunology1
ANTIBODY MOLECULES AND THE IMMUNE RESPONSE: THEORETICAL BACKGROUND
IAN M. ZITRON
Publisher Summary
The normal function of the immune system is to provide protection against invasion by pathogens. It is, unfortunately, also the source of less welcome events, such as graft rejection and autoimmune diseases, in which the system reacts against normal body constituents. An immune response, whether it be beneficial or harmful, is a multi-layered phenomenon. There are two forms of immunities present in a living body—innate immunity that is present through the course of the being’s lifetime and acquired immunity that stems from adaptations by the body to foreign stimulus with effector functions working in response to that particular stimulus. Adaptation shows itself in two characteristics, specificity and memory, that combine to give the system its efficiency. The three defining characteristics of the acquired response—specificity, memory, and self/nonself discrimination—reflect the properties and activities of lymphocytes. These cells are directly responsible for the acquired immune response, although in many instances the response also requires the involvement of cells of the monocyte/macrophage lineage.
I THE ORGANIZATION OF IMMUNE PROTECTION
The normal function of the immune system is to provide protection against invasion by pathogens. It is, unfortunately, also the source of less welcome events, such as graft rejection and autoimmune diseases, in which the system reacts against normal body constituents. An immune response, whether it be beneficial or harmful, is a multi-layered phenomenon. The purpose of this chapter is twofold. The first is to provide an introductory description of the various layers, in conjunction with an indication of some of the interactions that occur both between the layers and within them. The second is a more detailed consideration of those aspects of the system that are of most interest and importance to scientists who wish simply to use immunological techniques, particularly antibodies, in their research. The emphasis is on the genetic, cellular, and molecular events that give rise to the production of antibodies, rather than on immunochemistry per se. Nonimmunologists frequently complain that the language and terminology used in immunology present major barriers to an appreciation of the system, obscuring rather than clarifying. Although this chapter is in no sense a replacement for a comprehensive dictionary, it may make some aspects of the immune system more understandable.
II INNATE AND ACQUIRED IMMUNITY
The first broad separation is the discrimination between innate and acquired immunity. Innate immunity is, as its name suggests, present at birth and persists throughout life. It represents the first line of defense against insult (the word “insult” is used broadly in immunology and refers to invasion by pathogenic microorganisms, parasites, and, in some instances, tumors) and is composed of a number of physical, cellular, and chemical barriers. Skin and mucous membranes physically impede invasion. Chemical barriers include gastric pH, enzymes such as lysozyme in tears and saliva, and other biologically active molecules such as the interferons and the proteins of the complement system. Cells, such as polymorphonuclear leukocytes (PMNs) and natural killers (NKs) are also integral components of this aspect of protection. Thus at least three layers of protection exist in innate immunity. Although each is mechanistically quite different in the way in which protection is effected, there are characteristics that all layers have in common. Each layer is either continuously present, or very rapid in response to insult. The layers show no specificity vis a vis the insult: they are incapable of discriminating between different pathogens. Also, they show no memory of prior insults: neither the quality nor the quantity of innate immunity is increased by a second exposure to the same pathogen. Acquired immunity differs from innate immunity in each of these characteristics. Table I shows a comparison between innate and acquired immunity. The elements of innate immunity provide a significant degree of protection; the severity of immune deficiency syndromes that reflect a lack of individual components of this system provides ample evidence of its importance.
TABLE I
Differences between Innate and Acquired Immunity
| Property | Innate immunity | Acquired immunity |
| Components | Physical barriers, e.g., skin; chemical barriers, e.g., lysozyme, interferons; cellular components, e.g., NK cells | Lymphocytes and accessory cells |
| Specificity | None | Specific for insult or pathogen |
| Presence at birth | Yes | Some elements; others develop postnatally |
| Effect of exposure to insult | None; no memory generated | Effector function and memory elicited |
In contrast, acquired immunity is adaptive, since it responds to the particular insult with effector functions that act only on that insult. Adaptation shows itself in two characteristics, specificity and memory, which combine to give the system its efficiency. Figure 1 shows the time courses and magnitudes of (antibody) responses to two immunologically unrelated substances, designated A and B. The figure illustrates the difference in response subsequent to first (primary) and second (secondary) exposure to a foreign substance; this behavior reflects immunological memory. Specificity is the ability to discriminate between different insults; in the figure, the primary immunization with A elicits antibodies that recognize A only; the memory that this response generates is specific for A. Specificity may be maintained even when the substances are closely related biologically and chemically. One of the best known examples of the system’s ability to distinguish subtle differences is the human ABO blood group system (Landsteiner, 1946). Perhaps an even more striking example of the level of discrimination possible is that antibody populations may be identified that are capable of discriminating between closely related nitrophenyl groups (Little and Eisen, 1969). Immunological memory is very much like that observed in the functioning of the nervous system: the cells have the ability to recognize, and respond to, the second (or third, and so on) exposure to a given insult in a way that is both quantitatively and qualitatively different from the response to the first, or primary, exposure. Moreover, memory is itself specific because the enhanced response subsequent to re-exposure is directed only toward the foreign substance used for initial immunization.
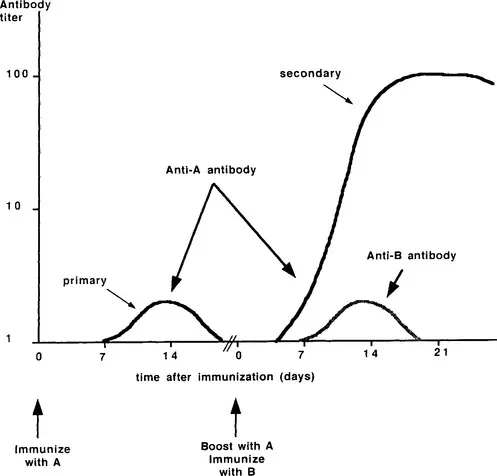
FIGURE 1 Kinetic and quantitative differences between primary and secondary humoral (antibody) responses. The primary response to immunization with A shows a lag period of ~7 days, followed by a logarithmic increase, a relatively low plateau titer, and a rapid decay. In contrast, secondary immunization with A gives rise to a response with a shorter lag period, a steeper logarithmic increase reaching a significantly higher plateau (note the logarithmic scale of the ordinate), and a slow decay phase. The difference between primary and secondary responses reflects immunological memory, which has been generated as a consequence of primary immunization, but which requires rechallenge to be expressed. The secondary immunization with A is accompanied by primary immunization with an immunologically distinct substance, B. The indicated response to B is characteristic of a primary response, indicating that memory itself is specific.
The acquired immune response shows an additional characteristic that is of considerable significance for homeostasis, the ability to discriminate between self and nonself. The phenomenon was first shown in the 19th century by Paul Ehrlich (1900), who gave it the name “Horror Autotoxicus.” In its simplest terms, the observation is that, although vertebrates may be immunized with foreign (i.e., nonself) material and be shown to mount an immune response, spontaneous responses to the animal’s own molecules leading to autoimmune disease (i.e., anti-self) are rare. The process by which recognition of self is minimized is referred to as self-tolerance. Self-tolerance is not simply a failure to recognize self components, but is an active process involving the regulation of lymphocyte survival and function (see subsequent text and Figures 7 and 8; Nossal, 1994).
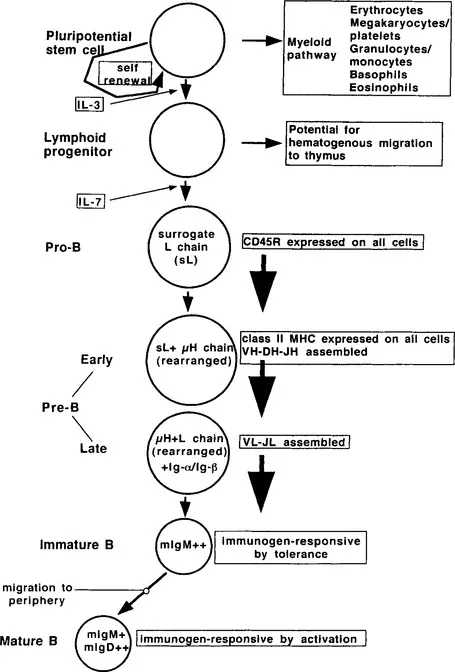
FIGURE 7 Developmental sequence of B cells. B cells are derived from the pluripotential stem cell, which has self-renewing capacity and is also able to give rise to all elements of the hemopoietic system. Interleukin-3 (IL-3) is required for development of the lymphoid progenitor, a cell that is irreversibly committed to the lymphocytic pathway. This cell can give rise to both B and T lymphocytes. The latter require migration from bone marrow to thymus for development. Pro-B cell development is driven by IL-7. Pro-B cells express CD45 and surrogate light chain and are the first unambiguous step in the B cell pathway. Pre-B cells are the stage at which class II MHC becomes expressed and the Ig loci undergo the rearrangements that result in the expression of variable region domains. The heavy chain locus rearranges prior to the light chain. Immature B cells express mIgM and the signal-transducing complex of Ig-α and Ig-β. As such, these are the first cells in the pathway to be responsive to external immunogen. However, exposure to immunogen results in tolerization. The final step is the mature B cell. This is recognized by the co-expression of mIgM and mIgD. These cells migrate to the...
Table of contents
- Cover image
- Title page
- Table of Contents
- Copyright
- CONTRIBUTORS
- PREFACE
- Chapter 1: ANTIBODY MOLECULES AND THE IMMUNE RESPONSE: THEORETICAL BACKGROUND
- Chapter 2: ADJUVANT-FREE POLYCLONAL ANTIBODY RESPONSE MANIPULATED BY ANTIBODY-MEDIATED ANTIGEN TARGETING
- Chapter 3: HYBRIDOMAS AND MONOCLONAL ANTIBODIES
- Chapter 4: PCR AMPLIFICATION OF ANTIBODY GENES
- Chapter 5: LABORATORY ANIMAL CARE AND USE
- Chapter 6: ADJUVANTS
- Chapter 7: LARGE-SCALE PRODUCTION OF MONOCLONAL ANTIBODIES
- Chapter 8: ANTIBODY-BINDING BACTERIAL PROTEINS AS IMMUNOREAGENTS
- Chapter 9: CONJUGATING ANTIBODIES TO LIGANDS, ENZYMES, AND METALLIC PARTICLES
- Chapter 10: DOT IMMUNOBINDING (DIB), ENZYME-LINKED IMMUNOSORBENT ASSAY (ELISA), AND RADIOIMMUNOASSAY (RIA) FOR DETECTING PEPTIDE ANTIGENS AND SPECIFIC ANTIBODIES
- Chapter 11: IMMUNOSTAINING CELLS AND TISSUES
- Chapter 12: PROTEIN IMMUNOBLOTTING
- Chapter 13: FLOW CYTOMETRY AND FLUORESCENCE-ACTIVATED CELL SORTING
- Chapter 14: SCREENING EXPRESSION LIBRARIES WITH ANTIBODIES
- Chapter 15: IMMUNOAFFINITY PURIFICATION
- INDEX
Frequently asked questions
Yes, you can cancel anytime from the Subscription tab in your account settings on the Perlego website. Your subscription will stay active until the end of your current billing period. Learn how to cancel your subscription
No, books cannot be downloaded as external files, such as PDFs, for use outside of Perlego. However, you can download books within the Perlego app for offline reading on mobile or tablet. Learn how to download books offline
Perlego offers two plans: Essential and Complete
- Essential is ideal for learners and professionals who enjoy exploring a wide range of subjects. Access the Essential Library with 800,000+ trusted titles and best-sellers across business, personal growth, and the humanities. Includes unlimited reading time and Standard Read Aloud voice.
- Complete: Perfect for advanced learners and researchers needing full, unrestricted access. Unlock 1.5M+ books across hundreds of subjects, including academic and specialized titles. The Complete Plan also includes advanced features like Premium Read Aloud and Research Assistant.
We are an online textbook subscription service, where you can get access to an entire online library for less than the price of a single book per month. With over 1.5 million books across 990+ topics, we’ve got you covered! Learn about our mission
Look out for the read-aloud symbol on your next book to see if you can listen to it. The read-aloud tool reads text aloud for you, highlighting the text as it is being read. You can pause it, speed it up and slow it down. Learn more about Read Aloud
Yes! You can use the Perlego app on both iOS and Android devices to read anytime, anywhere — even offline. Perfect for commutes or when you’re on the go.
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Please note we cannot support devices running on iOS 13 and Android 7 or earlier. Learn more about using the app
Yes, you can access Antibody Techniques by Vedpal S. Malik,Erik P. Lillehoj in PDF and/or ePUB format, as well as other popular books in Medicine & Immunology. We have over 1.5 million books available in our catalogue for you to explore.